Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механизмы инициации движенияСодержание книги
Поиск на нашем сайте
Сейчас уже многое известно о функциях моторной коры. Ее рассматривают как центральную структуру, управляющую самыми тонкими и точными произвольными движениями, посылающую свои сигналы к мотонейронам спинного мозга (так называемая пирамидная система). Именно в моторной коре строится конечный и конкретный вариант моторного управления движением. Моторная кора использует оба принципа управления: контроль через петли обратной сенсорной связи и через механизм программирования. Это достигается тем, что к ней сходятся сигналы от мышечной активности, от сенсомоторной, зрительной и других отделов коры, которые и используются для моторного контроля коррекции движения. Кроме того, к моторной коре приходят сигналы, связанные с программированием движения из передних отделов коры и подкорки. Каким образом в моторной коре совмещаются оба принципа управления? Чтобы ответить на этот вопрос, прежде всего необходимо рассмотреть механизмы, с помощью которых в мозге осуществляется актуализация моторных программ, хранящихся в ЦНС. Сейчас уже накопилось много данных о том, что моторная кора получает информацию из структур, имеющих отношение к хранению двигательных программ. Так, в опытах с регистрацией нейронной активности у обезьян было показано, что при выполнении ими заученного движения активность нейронов зубчатого ядра мозжечка на 10 мс опережает изменение активности нейрона в моторной коре, которое предшествует появлению мышечного движения. Значение сигналов, поступающих из мозжечка, для активности нейронов моторной коры было также установлено в опытах с временным понижением температуры мозжечка у обезьян. Исследователи нашли, что во время охлаждения мозжечка импульсация нейронов моторной коры и соответствующее выученное движение запаздывали. Кроме сигналов от мозжечка в моторную кору поступают сигналы от базальных ганглиев — структуры, которая ответственна за хранение главным образом двигательных программ врожденного поведения (пищевого, питьевого и др.). Показано, что клетки базальных ганглиев, так же как и мозжечка, разряжаются задолго до движений, совершаемых животными в ответ на сигнал. По данным Н.Ф. Суворова, разряды нейронов хвостатого ядра кошки на 50-150 мс опережают ЭМГ-компонент хватательного движения лапы, направленного на пищу, т. е. возникающего как натуральный пищевой рефлекс. В хвостатом ядре были найдены также нейроны, у которых возбуждение опережало условную двигательную пищевую и оборонительную реакции. Эти реакции у нейронов появлялись с выработкой условного рефлекса и исчезали с его угашением. Нейроны с реакцией, опережающей двигательные оборонительные и пищевые условные рефлексы, локализованы в различных частях хвостатого ядра.
Таким образом, моторная кора управляет движением, используя информацию, поступающую как по сенсорным путям от других отделов коры, так и от генерируемых в ЦНС моторных программ, которые актуализируются в базальных ганглиях и мозжечке и доходят до моторной коры через таламус, где они, по-видимому, предварительно взаимодействуют. Сигналы от мозжечка и базальных ганглиев распространяются не только вверх к коре, но и вниз, по нисходящему пути к спинному мозгу (так называемый экстрапирамидный путь). Однако само подразделение двигательной системы на пирамидную и экстрапирамидную, как считает П. Милнер, является источником путаницы и ошибок. Возможно, оно явилось следствием заблуждения, возникшего из начального представления о том, что пирамидная система, берущая начало от моторной коры, — единственная двигательная система. Поэтому те структуры мозга, роль которых в регуляции движения была установлена позже, были объединены в экстрапирамидную систему. Между этими системами трудно провести функциональную грань. И как видно на примере моторной коры, обе системы тесно взаимодействуют в процессе построения движения. Согласно гипотезе, высказанной П. Робертсом, актуализация моторных программ происходит через активацию командных нейронов. Сами командные нейроны могут контролироваться и затормаживаться сверху. Снятие торможения с командных нейронов повышает их возбудимость и тем самым высвобождает «предпрограммированные» цепи для той деятельности, для которой они предназначены. По-видимому, существует общий биохимический механизм актуализации моторных программ через рост активности ДА- и НА-ергических систем в мозге. Эти системы обнаруживают свойство тормозить тормозные интернейроны и тем самым растормаживать нейронные сети.
Имеются данные о наличии в стриатуме ГАМК-ергических тормозных интернейронов, которые тонически тормозят запрограммированные нейронные цепи, управляющие позой и движениями, а сами тормозятся ДА-ергическими нейронами из черной субстанции. Показано, что НА-волокна проецируются в мозжечок, их активация также ведет к торможению тормозных интернейронов. Сейчас многие исследователи разделяют точку зрения, что ДА-и НА-ергические пути участвуют в вытормаживании тормозных интернейронов в структурах, вносящих вклад в инициацию и двигательный контроль поведения. Таким образом, в базальных ганглиях, мозжечке уже заложен механизм, который может актуализировать хранящиеся там двигательные программы. Однако, для того чтобы привести в действие весь этот механизм, необходимо, чтобы в эти структуры поступил сигнал, который послужил бы начальным толчком для этого процесса. Изучение этого механизма привело к представлению о двух системах инициации движения. Одна из них — это лимбическая система мозга, по терминологии Ю. Конорского [14], «эмоциональный мозг».С помощью этой системы осуществляется «трансляция мотивации в действие», т. е. в действия, которые связаны с утолением голода, устранением страха и удовлетворением других потребностей. Согласно данным Г. Могенсона, рассматривающего механизмы инициации локомоции, эта трансляция достигается за счет особого пути передачи сигналов от лимбических структур к базальным ганглиям. При этом важным звеном, связывающим лимбическую систему с моторной, является прилегающее ядро (n.accumbens). Его возбуждение сигналами из лимбической системы, по-видимому, через механизм активации ДА-ергических путей, ведет к актуализации врожденных поведенческих программ. С появлением ассоциативной коры у высших животных увеличивается роль когнитивных процессов. Появляется вторая система инициации движения — «когнитивный мозг». Он обеспечивает инициацию различных специфических движений в соответствии с инструкцией, установкой на ответ, прошлым опытом и обучением. Приход сигнала от ассоциативной коры в неостриатум базальных ганглиев, который имеет обширные сенсорные проекции, растормаживает его командные нейроны и тем самым актуализирует двигательные программы, главным образом двигательные автоматизмы, выученные движения. Эти программы через таламус также достигают моторной коры. Таков путь, через который «когнитивные команды» инициируют программы движений, сохраняемые в ЦНС. Согласно Г. Могенсону, прилегающее ядро и хвостатое ядро фильтруют сигналы соответственно от лимбической системы («эмоционального мозга») и от церебральной коры («когнитивного мозга»). Хотя в процессе филогенетического развития роль «когнитивного мозга» в инициации двигательных ответов возрастает, «эмоциональный мозг» и «когнитивный мозг» обычно действуют совместно. На рис. 40 схематично представлена система связей различных структур мозга, по Г. Могенсону, через которые эмоциональные и когнитивные процессы транслируются в движение. Согласно схеме все процессы управления движением делятся на 3 фазы: блок инициации движения, включающий лимбическую систему с прилегающим ядром (n. accumbens), и ассоциативную кору, блок программирования движения, включающий мозжечок, базальные ганглии, моторную кору, таламус как посредника между ними, а также спинальные и стволовые генераторы; исполнительный блок, охватывающий мотонейроны и двигательные единицы. На рисунке обратные связи опущены. Но нужно иметь в виду, что управление движением включает наряду с командами по прямым связям интеграцию обширной проприоцептивной и экстероцептивной информации.
Эмоциональный мозг (лимбическая система) действует через n. accumbens и далее через базальные ганглии. Когнитивный мозг (ассоциативная кора) действует на
Рис. 40. Схема инициации двигательного акта «эмоциональными и когнитивными» процессами (по Г. Могенсону, 1977)
Схема, представленная на рис. 40, допускает объяснение не только инициации непроизвольных движений, врожденных форм двигательного поведения и автоматизмов, но и произвольных движений. Рефлексы и произвольные движения не противоречат друг другу. Произвольные движения также подчиняются рефлекторному принципу. Это особенно очевидно в случае оперантных или инструментальных реакций животных, которые являются простейшей формой произвольных движений. Что же отличает произвольное движение от непроизвольного? По определению шведского нейрофизиолога Р. Гранита, которое в 1977 г. он приводит в своей книге «Целенаправленный мозг», «произвольным в произвольном движении является его цель». Цели произвольных движений могут быть бесконечно разнообразными. В. Гурфинкель также определяет произвольное движение в связи с его целью. Рассматривая профессиональные движения стрелков-спортсменов — прицеливание, он отмечает, что особенностью хорошо управляемого движения у стрелка является точное удержание цели. Пистолет неподвижен, тогда как многие части тела двигаются, т.е. положение руки стрелка в пространстве стабилизируется всякого рода рефлекторными механизмами: вестибуло-окулярной системой, вестибуло-спинальной системой и др. Произвольные движения человека — это сознательно регулируемые движения. И в отличие от животных они являются специфически человеческой функцией и тесно связаны с речью. У человека цели произвольных движений формируются с помощью речевых процессов. Роль речевого опосредования в превращении непроизвольных импульсивных движений у детей в произвольные и сознательно управляемые была изучена А.Р. Лурией и его сотрудниками. Они показали, что активизация детской речи — предварительное планирование в речевой форме предпринимаемых действий — ведет ребенка к овладению своим поведением, тогда как задержка в речевом развитии у аномальных детей приводит к снижению уровня произвольной или волевой регуляции двигательного поведения ребенка.
Произвольные движения, вызываемые инструкцией или внутренним побуждением человека, опосредованы внутренней речью, претворяющей замысел (цель) во внутренний план действий. Произвольные движения так же, как и непроизвольные, являются результатом сопряженной работы «когнитивного» и «эмоционального» мозга. Таким образом, управление и контроль за движением — достаточно сложный процесс. Он включает обработку информации, получаемой через прямые и обратные связи между передними отделами коры, моторной корой, мозжечком, базальными ганглиями, а также стволом мозга и спинным мозгом. Важная роль принадлежит проприоцептивной и экстероцептивной афферентации. Двигательная система организована по иерархическому принципу с постепенным увеличением сложности сенсомоторной интеграции. Двигательная система работает как многоуровневая. На каждом уровне имеется своя «ведущая афферентация» и собственный тип регулируемых движений. Функциональные состояния Понятие функционального состояния (ФС) широко используется в психологии, физиологии, эргономике и т. д. Это объясняется тем, что в самых различных сферах деятельности человека успешность его труда, обучения, творчества, физическое и психическое здоровье в значительной мере зависят от его ФС. Отношения ФС и эффективности выполняемой работы принято описывать в виде куполообразной кривой. Тем самым вводится понятие оптимального функционального состояния, при котором человек достигает наиболее высоких результатов. Поэтому управление ФС является одним из важных резервов, который может быть использован для повышения эффективности деятельности человека на производстве, в школе, в вузе и в других сферах общественной практики. Оптимизация ФС является непременным условием формирования здорового образа жизни. Однако на сегодняшний день, несмотря на очевидность практической значимости проблемы ФС, методы диагностики и оптимизации ФС остаются недостаточно изученными. В значительной мере такая ситуация обусловлена неразработанностью теории ФС и отсутствием четкого понятийного аппарата. Это касается и самого понятия ФС. Наиболее часто ФС определяют как фоновую активность ЦНС, в условиях которой осуществляется та или иная деятельность. При этом И.П. Павлов говорил о тонусе коры больших полушарий или ее возбудимости. Позже работами П.К. Купалова показано, что изменения ФС отражаются не только в возбудимости головного мозга, но и в его реактивности и лабильности. Очевидно, что такое определение ФС носит самый общий характер и требует конкретизации. Шаги, сделанные исследователями в этом направлении, привели к выводу, что существует большой набор физиологических реакций организма, в которых отражаются сдвиги фоновой активности ЦНС. Это ЧСС, АД, частота и глубина дыхания, скоростные характеристики двигательных реакций, изменения ЭЭГ и т. д.
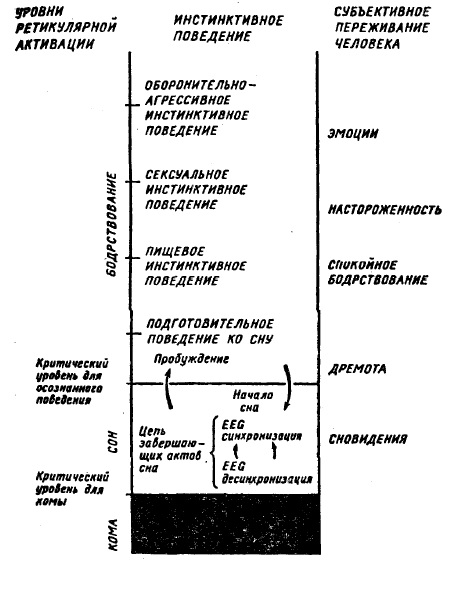
Все это побудило исследователей, уточняя определение ФС, обратиться к описанию тех поведенческих реакций, в которых проявляются его изменения. Наиболее широкое распространение при этом получили два подхода. При первом ФС определяется через комплекс взаимосвязанных физиологических реакций. Иначе говоря, изменения ФС — это смена одного комплекса реакций другим. В свете таких представлений диагностика и идентификация ФС рассматриваются как решение задачи распознавания многомерного вектора, компонентами которого являются различные физиологические показатели или реакции. Поиск и нахождение все новых и новых реакций и их комбинаций, в которых отражается динамика ФС, имеет под собой разумную основу, так как, по-видимому, существуют уникальные комбинации реакций, соответствующие определенным ФС. Кроме того, с увеличением числа регистрируемых физиологических параметров достигается более дробное представление о тех процессах, которые характеризуют изучаемое конкретное ФС. Однако определение ФС через перечень различных реакций, найденных эмпирическим путем, имеет свои ограничения, так как остается невыясненным их отношение к реальным механизмам формирования функциональных состояний. Второй подход наиболее популярен в эргономике, среди инженерных психологов. Он основывается на данных зависимости эффективности условнорефлекторной деятельности животных, работоспособности и результатов деятельности человека от ФС. При этом ФС оценивается не столько по физиологическим реакциям, сколько по результатам трудовой деятельности человека, которые рассматриваются как наиболее интегральный показатель ФС. Снижение эффективности выполняемой работы расценивается как признак ухудшения ФС. Дальнейшее развитие этого подхода выразилось в создании психометрических тестов, включающих решение различных когнитивных задач. Конечно, располагая данными о количестве и характере допущенных ошибок, о сбое в работе, можно судить о степени выраженности тех или иных экстремальных состояний, возникающих в результате утомления, монотонии и т. п. Такой способ оценки ФС безусловно полезен для практики, так как для конкретных видов трудовой деятельности позволяет решать многие задачи повышения эффективности труда и предотвращать развитие у человека нежелательных, недопустимых ФС. Вместе с тем этот метод в основном лишен прогностической силы, так как он способен выявлять только наиболее явные, грубые изменения в ФС. Определение ФС через комплексы поведенческих, физиологических реакций или успешность производственных операций, выполняемых человеком, в лучшем случае дает возможность обнаружить изменения в состоянии, но ничего не говорит о том, что есть само функциональное состояние, так как не касается нейрофизиологических механизмов регуляции ФС. При таком подходе, отмечает Е.П. Ильин, происходит подмена определения сущности состояния описанием сдвигов, происходящих при его изменении. Одновременно предполагается молчаливое отрицание наличия самостоятельного нейрофизиологического механизма с функцией регуляции ФС. Однако такое представление противоречит многочисленным нейрофизиологическим данным. Исследование модулирующих систем мозга: ретикулярной формации с ее активирующими и инактивирующими отделами, а также лимбической системы, от которой зависит мотивационное возбуждение, дает основание выделять их в особую функциональную систему, которая имеет несколько уровней реагирования: физиологический, поведенческий и психологический (субъективный). Выражением активности этой функциональной системы и является ФС. Функциональное состояние — психофизиологическое явление со своими закономерностями, которые заложены в архитектуре особой функциональной системы. Такой взгляд на ФС подчеркивает важность изучения собственных механизмов регуляции ФС. Только на основе знания о реальных процессах управления ФС можно создавать адекватные методы диагностики ФС, как наиболее отвечающие его основным закономерностям. Определение ФС через поведенческие реакции приводит к отождествлению ФС с понятием уровня бодрствования. Предложение отделить понятие «уровня бодрствования» от понятия «уровень активности» нервных центров (функционального состояния) впервые было высказано В. Блоком. Уровень бодрствования рассматривается им как поведенческое проявление различных уровней функционального состояния. Он допустил, что между сном и крайним возбуждением существует непрерывный ряд изменений уровня бодрствования, с которым монотонно связаны изменения в уровне активации нервных центров. Идея о том, что уровень активации нервных центров обусловливает уровень бодрствования, легла в основу и схемы Дж. Моруцци (рис. 41). Согласно его представлениям разные формы инстинктивного поведения, включая сон, могут быть размещены на шкале уровней бодрствования. Каждому типу инстинктивного поведения соответствует определенный уровень ретикулярной активации. В отличие от В. Блока субъективные переживания рассматриваются Дж. Моруцци как составная часть каждого инстинктивного поведения и размещаются на шкале уровней бодрствования. В схемах В. Блока и Дж. Моруцци ни для функционального состояния, ни для уровня бодрствования не вводится количественной меры. Экспериментально соотношение уровня бодрствования и ФС изучалось E.H. Соколовым и H.H. Даниловой. В качестве предполагаемой меры уровня функционального состояния была взята средняя частота спайковых разрядов неспецифических и специфических нейронов таламуса кролика. Уровень бодрствования оценивался по комплексу ЭЭГ, вегетативных (дыхательных) и моторных реакций. Было найдено, что ранг уровня бодрствования линейно зависит от функционального состояния мозга, оцениваемого средней частотой спайковых разрядов неспецифических нейронов таламуса кролика (рис. 42). В схеме, суммирующей полученные результаты и представления авторов о соотношении функциональных состояний, уровней бодрствования и инстинктивного поведения (безусловных рефлексов) с эффективностью исполнения задания, классификация инстинктивного поведения, предложенная Дж. Моруцци, дополнена ориентировочным поведением. Безусловные рефлексы: оборонительный, пищевой, половой, ориентировочный, переход ко сну, сон — располагаются
Рис. 41. Соотношения уровней рекулярной активации, форм инстинктивного поведения и субъективных переживаний. Эмоции представлены в качестве одного из видов эмоционального переживания. Каждому уровню ретикулярной активации соответствует определенный тип инстинктивного поведения. Выделена подготовка ко сну как форма поведения (по ж. Моруцци, 1969)
Рис. 42. Зависимость между функциональным состоянием, уровнем бодрствования, инстинктивным поведением Рис. 42. Зависимость между функциональным состоянием, уровнем бодрствования, инстинктивным поведением (безусловными рефлексами) и эффективностью выполнения задания (по Е.Л. Соколову, H.H. Даниловой, 1974). Виден линейный характер зависимости между частотой разрядов нейрона и уровнем бодрствования или неспецифическим компонентом поведения разного типа и нарушение линейной зависимости для общей эффективности; эмоции здесь не представлены, они размещены вдоль всей оси уровня бодрствования
по шкале уровней бодрствования и каждому из них соответствует определенный уровень функционального состояния. В этой схеме функциональное состояние выделено в самостоятельное явление. В последнее время существенно уточняются функции модулирующих систем и, следовательно, механизмов регуляции ФС. При этом выявлена их большая значимость для поведения, чем это представлялось ранее. Взгляд на ФС лишь как на фактор, ухудшающий или улучшающий выполнение деятельности, сменился представлением о его более фундаментальной роли в поведении. Так, показано, что снижение уровня активации нервной системы у крыс, в частности за счет истощения в ЦНС дофамина (ДА), полностью нарушает как приобретенные при жизни навыки, так и врожденное мотивированное поведение. Если такую крысу бросить в бассейн с водой, то она погибает, так как не может плыть. Однако если ее ущипнуть за хвост или охладить воду в бассейне, что действует стимулирующим образом на животное, то крыса плывет и спасается. Таким образом, сохранность нервных связей, определяющих паттерн конкретного поведения, еще не гарантирует его реализацию. Другой пример может быть взят из работ, в которых изучались природные и приобретенные свойства у нейронов-детекторов. Известно, что свойства детекторов зрительной коры, хотя в основном генетически детерминированы, но тем не менее окончательно они формируются в сенситивный период и сильно зависят от стимульного окружения, в котором содержится новорожденное животное. Так, у котят, воспитанных в окружении вертикальных линий (в вертикальной среде), нейроны зрительной коры реагируют преимущественно на вертикально-ориентированные полосы, тогда как большая часть зрительных нейронов котят, воспитанных в горизонтальной среде, реагирует на горизонтальные линии. Особенности зрительного воспитания впоследствии сказываются и на поведении. Воспитание в горизонтальной среде вызывает у котенка трудности при передвижении между ножками стула. Котята, содержащиеся в вертикальной среде, затрудняются при хождении по ступенькам лестницы. В опытах В. Зингера у котят в сенситивный период изменяли бинокулярную стимуляцию глаз. Для этого один глаз закрывали или изменяли его положение в орбите (искусственное косоглазие). В результате у нейронов-детекторов зрительной коры формировались аномальные связи: детекторы теряли свойство бинокулярности. Далее было показано, что модификация свойств у детекторов в сенситивный период под влиянием сенсорной среды возможна лишь при сохранении модулирующего, активирующего входа к ним от ретикулярной формации среднего мозга и таламической неспецифической системы. Если устранить эти неспецифические, активирующие влияния, например разрушением ретикулярного ядра таламуса, что сопровождалось потерей ориентировочной реакции на стимул, то процесс формирования детекторов в сенситивный период приостанавливался в том полушарии, связи которого с ретикулярным ядром таламуса разрушались, и сохранялся в другом полушарии, которое продолжало получать модулирующие, неспецифические воздействия от своего ретикулярного ядра. Таким образом, для нормального формирования в сенситивном периоде свойств у нейронов-детекторов необходимы ориентировочные реакции, которые предполагают активацию от модулирующей неспецифической системы мозга. Модулирующие влияния в ЦНС играют не менее важную роль для процесса обучения, чем, скажем, подкрепление условного раздражителя безусловным. Показано, что примерно у 40% нейронов гиппокампа кролика можно выработать условные рефлексы при сочетании звукового стимула с электрокожным раздражителем. При этом параллельно со становлением условного рефлекса идет процесс формирования другой условнорефлекторной связи, специально изменяющей состояние данного нейрона, что выражается в росте его фоновой активности — частоты спайков («ассоциативный тонический ответ»). Если по каким-либо причинам условнорефлекторное изменение состояния данного нейрона не возникает, то и вырабатываемый у него условный рефлекс не обнаруживается. Это дало основание для заключения, что ассоциативный процесс включает в себя формирование состояния, качественно специфического для каждой временной связи. Данное явление — один из ведущих механизмов формирования условнорефлекторного поведения. Деятельность не существует отдельно от состояния и выступает как единое целое с ним. Таким образом, существует два механизма условнорефлекторной деятельности: 1) настроечный, регулирующий состояние мозга и создающий определенный уровень возбудимости и работоспособности нервных центров и 2) запускающий, который инициирует ту или иную условную реакцию. Б.И. Котляр вводит принцип функционального полиморфизма мозга, согласно которому состояние мозга соответствует виду деятельности и каждому состоянию соответствует качественная специфичность структуры церебральной нервной сети. Таким образом, механизм регуляции ФС является базальным механизмом интегративной деятельности мозга. От того, как функционирует модулирующая система мозга, зависит и обучение, и осуществление врожденного поведения. ФС в отношении поведения выполняет прежде всего операционную функцию. Оно включено в состав целенаправленного поведения как средство для его успешного осуществления. Вместе с тем ФС может выступать в качестве цели поведения. Это особенно очевидно выступает при рассмотрении ориентировочно-исследовательской деятельности и поведения регуляции цикла бодрствование—сон. Дефицит активации организма, особенно в условиях сенсорной депривации, побуждает человека и животных к ориентировочно-исследовательскому поведению. Многочисленные наблюдения свидетельствуют о стремлении животных к новому, к исследованию незнакомых пространств, к манипулированию сложными устройствами. О существовании самостоятельной потребности в новизне свидетельствует и то обстоятельство, что на ориентировочном подкреплении возможно обучение: выработка сложных инструментальных рефлексов у крыс, тонкой дифференцировки стимулов у обезьян и др. Другим типом поведения, в котором ФС выступает в качестве цели поведения, является сон. Согласно Дж. Моруцци, переход ко сну представляет определенный вид мотивированного поведения (инстинкта), в котором можно выделить подготовительные действия, завершающие акты и цель поведения. Цикл бодрствования—сон, с точки зрения Т.Н. Ониани, отражает циклическое чередование мотивации сна и мотивации бодрствования. В его основе работа гомеостатического механизма, реализуемого в смене и чередовании двух форм поведения, принципиальное отличие которых выражается в различии функциональных состояний: сна и бодрствования. Конкретное ФС человека и животного всегда зависит от целого ряда факторов. Это прежде всего мотивация, то, ради чего выполняется конкретная деятельность. Чем значимее мотивы, тем выше уровень ФС. Содержание самого труда — наиболее важный регулятор ФС. Уже в самом задании заложены определенные требования к специфике и уровню ФС. Общий уровень сенсорной нагрузки от сенсорного пресыщения до сенсорной депривации также изменяет ФС. Конкретное ФС зависит от исходного уровня активности нервной системы, сохраняющей след от предшествующей деятельности субъекта. Наконец, специфика и уровень ФС существенно зависят от индивидуальных особенностей субъекта, в частности от таких его свойств, как сила—слабость нервной системы, экстроверсия—интроверсия, тревожность и т. д. Так, индивиды с сильной нервной системой менее устойчивы к монотонии и раньше слабых показывают падение уровня активности нервной системы.
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-12-15; просмотров: 518; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.139.236.174 (0.02 с.) |

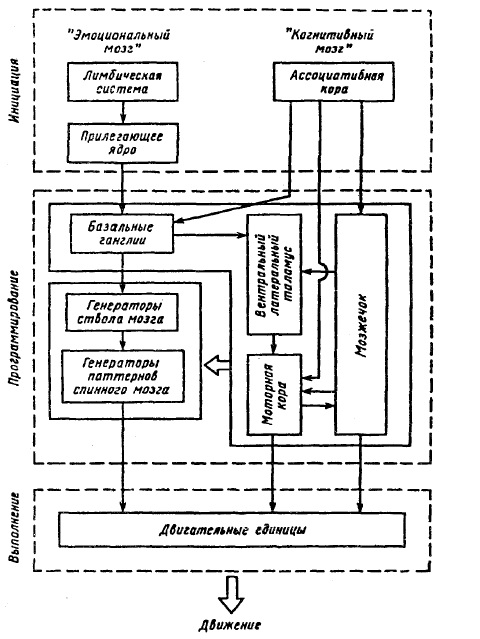 хвостатое ядро базальных ганглиев и параллельно на мозжечок. Моторная кора получает сигналы от мозжечка и базальных ганглиев через таламус.
хвостатое ядро базальных ганглиев и параллельно на мозжечок. Моторная кора получает сигналы от мозжечка и базальных ганглиев через таламус.