Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Общие свойства различных видов мотивацииСодержание книги
Поиск на нашем сайте
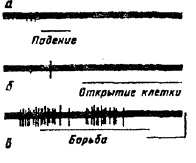
Несмотря на то, что виды мотивации различаются своей направленностью и способами удовлетворения потребности, всем им присущи некоторые общие черты. 1. Во время любой мотивации наблюдается активация моторной системы, хотя разные формы мотивации реализуются в разных паттернах двигательного возбуждения. Увеличивается средний уровень двигательной активности. Возрастает двигательная реактивность на сенсорные стимулы. Индифферентные раздражители приобретают способность стимулировать животное к движению. Возрастание моторной активности свойственно голоду, ярости и страху, любопытству. Единственным исключением, по-видимому, является пассивный страх, когда животное замирает на месте. 2. Повышение тонуса симпатической системы, выражающееся в вегетативных реакциях: росте частоты сердцебиения, артериального давления, возникновении сосудистых реакций, изменении проводимости кожи. Активация симпатической системы расширяет сосуды в скелетных мышцах, что обеспечивает увеличение притока кислорода к работающим мышцам. Повышение симпатического тонуса осуществляется через нисходящие пути, идущие от лимбической системы, гипоталамуса. 3. Рост активации афферентных систем, позволяющий животному с большей легкостью выявлять биологически значимые и сигнальные раздражители в окружающей среде. Это выражается в снижении сенсорных порогов, в усилении ориентировочных реакций. В ЭЭГ появляется десинхронизация как отражение усиления активирующих влияний, восходящих к коре от активирующей ретикулярной системы. 4. Возрастание поисковой активности (II фаза мотивации), которая носит целенаправленный характер. 5. Актуализация памяти является необходимым звеном для реализации поискового целенаправленного поведения, прежде всего образов цели и возможных способов их достижения. 6. Изменения в ЭЭГ, в которых отражается специфика мотивационного состояния. 7. Непременным проявлением мотивации является возникновение субъективных эмоциональных переживаний. Несмотря на то, что субъективные переживания страха, голода и других состояний различаются, их объединяет то, что все они относятся к переживанию с отрицательным эмоциональным знаком. Отрицательный эмоциональный тон — наиболее типичная форма субъективного отражения мотивации. Вместе с тем существует и другой класс субъективных переживаний с положительным знаком, который характеризует особую группу мотиваций: поведение любознательности, стремление к творческой активности. Мотивация как доминанта Мотивационное возбуждение, побуждающее к определенному целенаправленному поведению, обнаруживает свойство инертности. Оно длительно сохраняется, пока не будет удовлетворена породившая его потребность. Все посторонние раздражители только усиливают его, тогда как все другие виды деятельности подавляются мотивацией, которая реализуется в данном поведении. Мотивационное возбуждение, которое реализуется в поведении, получило название доминирующей мотивации. По этим свойствам мотивационное возбуждение тождественно явлению доминанты A.A. Ухтомского. Сходство мотивационного возбуждения с доминантой проявляется не только на поведенческом уровне. Применение нейрофизиологических методов показало, что во время мотивационного возбуждения и доминанты наблюдаются сходные изменения как в электрической активности мозга, так и в реакциях нейронов. Наиболее перспективным оказалось изучение моделей доминанты, создаваемых электрической или химической стимуляцией структур мозга. B.C. Русиновым при создании искусственной доминанты. использовалась анодная поляризация поверхности коры или воздействия на нее пульсирующим током. В этих опытах электрод с пульсирующим током обычно прикладывался к моторной коре. Сила тока была подпороговой и не вызывала движения конечности. Однако если к этой стимуляции добавить подпороговое болевое раздражение, то можно обнаружить, что пульсирующий подпороговый ток приобретает способность вызывать двигательные реакции животного, которые следуют в его ритме. Пульсирующие движения сохраняются в течение некоторого времени и после отключения токового раздражения. Это указывает, что в моторной коре создан очаг повышенной возбудимости со всеми свойствами доминанты (инертностью и способностью усиливаться за счет других возбуждений). В случае, когда доминанта в моторной коре создается с помощью анодной поляризации, показателем ее сформированности служит появление двигательных реакций конечности животного на индифферентные стимулы (звук, свет). С возникновением искусственной доминанты растет негативность постоянного потенциала, регистрируемого от коры больших полушарий. Аналогичное изменение постоянного потенциала коры возникает во время ЭЭГ-реакции активации. Так его можно зарегистрировать от коры животного во время ориентировочного рефлекса, а также в ответ на электрическую стимуляцию активирующей ретикулярной формации среднего мозга и неспецифического таламуса. Возрастание негативности постоянного потенциала (ПП) можно получить и стимуляцией мотивационных Центров гипоталамуса, вызывающей у животных мотивационное целенаправленное поведение. Таким образом, сходное возрастание негативности ПП во время искусственной доминанты и мотивации указывает на повышение возбудимости структур мозга — свойство, которое, по A.A. Ухтомскому, характерно для каждой естественной доминанты. Сама анодная поляризация коры меняет не только фоновую активность, но и реакции нейронов на стимулы. У ранее ареактивных нейронов в ответ на световые раздражения возникают реакции учащения спайков, увеличивается интенсивность реакции у ранее реагировавших нейронов, тормозные реакции нейронов сменяются на возбудительные. Аналогичные изменения нейронов коры наблюдаются при электрической стимуляции мотивационных центров гипоталамуса. По данным Ю.Н. Хаютина, электрическая стимуляция центра голода в гипоталамусе сопровождается ростом лабильности у нейронов зрительной коры. Они начинали воспроизводить своими ответами более высокую частоту световых мельканий, чем раньше. Таким образом, у нейронов, охваченных мотивационным возбуждением, так же как и находящихся в центре доминанты, растет возбудимость и лабильность. Показано, что доминирующее мотивационное возбуждение существенно изменяет интегративные свойства нейронов различных структур мозга и прежде всего их конвергентные способности. Под влиянием электрической стимуляции гипоталамического центра голода, инициирующей мотивационное пищевое поведение, большинство нейронов сенсомоторной коры кролика, ранее не реагировавшие на световые, звуковые, гуморальные, а также биологически значимые раздражения, приобретали способность реагировать на эти раздражения. Только небольшое число нейронов не изменяет конвергентные свойства или даже снижает их. Анодическая поляризация латерального гипоталамуса вызывает противоположный эффект — ухудшение конвергентных свойств нейронов. Это говорит о том, что конвергентные свойства нейронов коры в значительной степени зависят от восходящих активирующих влияний гипоталамуса. Сходные изменения у нейронов обнаружены и при искусственной доминанте. Усиление конвергентных способностей нейронов объясняет такие свойства доминанты, как ее повышенная возбудимость и способность суммировать приходящие возбуждения. Доминирующая мотивация сходна с доминантой также тем, что она имеет в своей основе возбуждение некоторой функциональной констелляции центров, расположенных на различных уровнях центральной нервной системы. К.В. Судаков выделяет корково-подкорковый уровень интеграции мотивационного возбуждения. В этой интеграции одни структуры избирательно возбуждены, другие заторможены. При пищевой мотивации кролика, вызванной электрической стимуляцией так называемого центра голода латерального гипоталамуса, по данным К.В. Судакова, передние отделы сенсомоторной коры, где обычно и локализована голодная ЭЭГ-активация, оказывают тормозное влияние на инициативные центры голода латерального гипоталамуса, тогда как затылочная кора и ретикулярная формация среднего мозга, наоборот, активируют гипоталамический центр голода. При оборонительной мотивации у кролика, вызванной электрическим раздражением вентромедиального гипоталамуса, сенсомоторные и затылочные отделы коры, в которых появляется тета-ритм, как ЭЭГ-паттерн оборонительного возбуждения, также оказывают тормозные влияния на свой инициативный мотивационный центр гипоталамуса. Интегративный корково-подкорковый комплекс Мотивационного возбуждения активируется либо метаболической потребностью, либо специальными (ключевыми) стимулами, а у человека и социально значимыми. На уровне нейронов интеграция различных образований мозга в единую систему определенной биологической мотивации проявляется в появлении у нейронов общего единого ритма. По данным К.В. Судакова, во время различных форм мотивации (пищевой, оборонительной) у многих нейронов в самых разных структурах мозга регистрируется ритмическая активность в виде пачек спайков, регулярно следующих с интервалом около 150 мс. Пачечный тип активности с одним общим ритмом у многих нейронов рассматривается как механизм установлении межцентральных связей. Согласно A.A. Ухтомскому, усвоение единого ритма нервными центрами (принцип изолабильности) является механизмом их объединения в единую функциональную констелляцию. Эта точка зрения разделяется многими исследователями. Значительный вклад в развитие идеи A.A. Ухтомского об изолабильности как основном механизме установления корковых взаимосвязей внес М.Н. Ливанов. Им было показано существование положительной корреляции между скоростью выработки условного двигательного рефлекса на свет и степенью синхронности ритмических потенциалов в зрительной и двигательной корковых областях, т. е. в структурах, заинтересованных в формировании данного условного рефлекса. Синхронизация потенциалов различных структур мозга кролика увеличивается и при формировании временных связей, особенно на начальных этапах этого процесса. М.Н. Ливанов указывает на особую роль в этом процессе тета-ритма. Возрастание синхронизации электрической активности коры и подкорки было отмечено для ориентировочного рефлекса. Электрическое раздражение активирующей ретикулярной формации и гипоталамуса также порождает высокую синхронизацию электрической активности коры и подкорковых образований, а также различных областей коры головного мозга на частоте тета-ритма у кролика. Со сном синхронизация в электрической активности коры и подкорковых структур падает. Взаимодействие различных видов поведения строится на основе открытого A.A. Ухтомским принципа доминанты. В каждый данный момент времени деятельность организма определяется доминирующей в плане выживания и адаптации мотивацией. После завершения одного мотивированного поведения организмом завладевает следующая ведущая по социальной и биологической значимости мотивация. Ведущая мотивация подчиняет себе все другие. При формировании доминирующего поведения внутреннее состояние животного и человека и внешние стимулы нередко могут приходить в столкновение. Наиболее отчетливо это выступило при изучении роли обстановочной афферентации. В опытах X. Дельгадо с телеметрическим управлением поведением животных посредством электрической стимуляции через вживленные в мозг электроды показано, что поведение, вызываемое электрическим раздражением структур мозга, зависит также и от той среды, в которой содержится животное. В лабораторных условиях электрическая стимуляция латерального гипоталамуса обычно вызывает стандартную пищевую реакцию (поедание пищи) даже у сытого животного. Однако эта же стимуляция у обезьяны, содержащейся среди своих сородичей, может вызвать совсем другое поведение: оборонительное или половое. И это зависит от того, какое поведение демонстрируют другие особи стада. т. е. возбуждение, возникающее на обстановку, может оказаться более сильным, нежели то, которое вызывается электрической стимуляцией «центра голода». Доминирует и усиливается за счет других очагов возбуждения либо возбуждение, вызванное электрической стимуляцией мотивационного центра, либо возбуждение от внешних раздражителей. Возбуждение, которое станет доминирующим, и определит, какое поведение будет осуществлено. Нейроанатомия мотивации Эксперименты на животных с электрической стимуляцией и разрушением ядер гипоталамуса показали, что регуляция пищевой, питьевой, оборонительной мотивации осуществляется взаимодействием латерального и вентромедиального отделов в задней области гипоталамуса. Удобный объект наблюдения — коза. Сытая коза — спокойна, апатична. При раздражении латерального гипоталамуса (центра голода) возникает двигательное беспокойство и поисковое поведение: животное осматривается, хватает за одежду стоящих поблизости людей. При появлении пищи жадно ее ест. Но стоит убрать корм, вновь проявляет двигательное беспокойство, поисковое поведение. С прекращением раздражения животное вновь погружается в состояние апатии. При раздражении вентромедиального гипоталамуса, где расположен «центр насыщения», давно не кормленная, голодная коза становится апатичной, перестает есть, отворачивается от пищи, ложится. По прекращении раздражения животное сразу же возвращается к еде. Опыты с разрушением структур гипоталамуса проводились главным образом на крысах. При повреждении латерального гипоталамуса возникает поведение сытости. Крыса теряет интерес к еде, и, как следствие этого, — афагия, хотя исполнительный пищевой рефлекс сохраняется. Если пищу положить в рот, животное прожует ее и проглотит. Электрическое повреждение вентромедиального гипоталамуса приводит к гиперфагии: животное начинает есть больше, чем до операции, и прибавляет в весе. Со временем теория регуляции мотивации, базирующаяся на представлении о реципрокных взаимоотношениях двух центров, была поставлена под сомнение, так как основные результаты о существовании системы двух центров были получены с помощью методов электрической стимуляции и разрушения структур гипоталамуса. В этих случаях не исключается, что вызываемые эффекты обусловлены не столько воздействием на изучаемую структуру, сколько на проходящие через нее волокна. Однако изучение нейронной активности гипоталамуса, а также создание новых методов разрушения только клеточных структур при сохранности проводящих путей позволили реабилитировать мотивационную теорию двух центров. Дж. Гроссман и Я. Гроссман вводили в латеральный гипоталамус каиновую кислоту, которая избирательно разрушала тела нейронов и оставляла интактными нервные волокна. Они получили у животного афагию, что подтвердило концепцию о существовании в латеральном гипоталамусе центра, контролирующего голод. Недавно Ю. Оомура с сотрудниками применили другую кислоту (ибаиновую, которая менее токсична, чем каиновая) для разрушения тел клеток в вентромедиальном таламусе и смогли получить гипертрофию и ожирение у крыс. Это еще более подкрепило идею существования в гипоталамусе двух центров, контролирующих пищевое поведение. Нейроны двух областей гипоталамуса одинаково нужны для нормального пищевого поведения. По данным Ю. Оомура, при высокой пищевой мотивации сильно активизируются нейроны латерального гипоталамуса, а при насыщении — вентромедиального гипоталамуса. В латеральном гипоталамусе выявлены глюкозочувствительные нейроны, которые в ответ на электро-форетическое подведение к ним глюкозы, пропорционально дозе, тормозят свою активность без изменения сопротивления мембраны. В вентромедиальном гипоталамусе найдены глюкозорецептивные нейроны. На глюкозу они реагируют противоположно — в соответствии с величиной дозы увеличивают частоту разрядов, при этом сопротивление мембраны нейронов растет. Противоположные изменения активности нейронов глюкозорецептивной и глюкозочувствительной систем являются прямым доказательством того, что пищевая мотивация на уровне гипоталамуса регулируется реципрокными механизмами двух центров в латеральном и вентромедиальном отделах гипоталамуса. Пищевое поведение регулируется гуморальными и нервными механизмами. Их интеграция осуществляется в хемочувствительных нейронах гипоталамуса: глюкозочувствительных и глюкозореактивных нейронах. Эти нейроны обладают мультимодальной чувствительностью: к глюкозе, сахарным кислотам, свободным жирным кислотам, катехоламину, опиатам, инсулину, глюкогену и другим эндогенным химическим соединениям, кроме того, они реагируют на внешние сенсорные раздражения. Глкюкозочувствительные нейроны латерального гипоталамуса у обезьяны реагируют учащением спайков на вид и запах пищи, на вид шприца с глюкозой, на скорлупу ореха. Их активность модулируется пищевой мотивацией. Реакции этих нейронов на натуральные пищевые стимулы могут быть зарегистрированы только у голодной обезьяны. Введение глюкозы снижает их реактивность. По данным Е. Роллса, реакция нейронов, реагирующих на вид пищи (через 15-200 мс), предшествует реакциям других нейронов, которые реагируют на прием пищи (300 мс). При этом обнаружены нейроны, реагирующие только на вид пищи или только на вид воды. Глюкозочувствительные нейроны латерального гипоталамуса вовлекаются в процессе пищедобывательного поведения. У обезьян их активность резко возрастает за 2-0,8 с перед нажатием на рычаг для получения пищи и подавляется во время движения и следующего за ним подкрепления. Эти нейроны одновременно чувствительны к НА и опиатам. Помимо глюкозочувствительных нейронов в латеральном гипоталамусе обнаружены глюкозонечувствительные нейроны. Они активируются во время пищедобывательного поведения, однако их реакции совпадают с движениями, а не предшествуют им. По Ю. Оомура, эти нейроны также распознают ключевые экстероцептивные раздражители и принимают участие в реализации движений. Они чувствительны к ДА. Полагают, что глюкозонечувствительные нейроны играют роль в интеграции внешней информации, тогда как глюкозочувствительные нейроны участвуют в интеграции внутренней информации и восприятии подкрепления. Сейчас можно считать установленным, что реакции агрессивно-оборонительного типа также регулируются взаимодействием вентромедиальной и латеральной областей гипоталамуса через центральное серое вещество среднего мозга. Его повреждение либо сильно повышает порог электрического раздражения гипоталамуса для вызова реакции ярости, либо полностью блокирует ее. При электрическом раздражении Центрального серого вещества развивается сильно выраженная реакция ярости. Д. Адамсом в этой структуре обнаружены командные нейроны, запускающие реакцию агрессии и не возбуждающиеся при других реакциях (рис. 33). Исследователи говорят о различных видах агрессивности в зависимости от ее направленности на борьбу за самку, за место в групповой иерархии, за территорию, за самосохранение. Предполагается, что реализация этих разных форм агрессивного поведения обеспечивается различными участками в гипоталамусе. Поведение бодрствование—сон также обеспечивается системой двух центров. Уже К. Экономо на основе клинических наблюдений больных с повреждениями различных участков гипоталамуса предположил, что центр бодрствования локализован в заднем, а центр сна в его переднем отделах. С. Рэнсон, производя локальные повреждения различных участков гипоталамуса, подтвердил это мнение. В настоящее время считают, что гипоталамус является критической зоной для регулирования цикла бодрствование—сон. Это мнение подтверждается и тем, что как высокочастотное, так и низкочастотное электрическое раздражение преоптической области гипоталамуса вызывает синхронизацию Рис. 33. Реакция нейрона «агрессии» в центральном сером веществе; а — клетка молчит, когда кошку поднимают и роняют на пол; б — нейрон разряжается только один раз, когда между кошками нет преграды, но борьбы нет; в — нейрон генерирует частые спайки во время драки кошек; калибровка: 100 мкв, 1 мс (по Д.Адамсу, 1968)
электроэнцефалограммы и поведенческий сон. Противоположный эффект — поведенческое и электроэнцефалографическое пробуждение Т.Н. Ониани наблюдал при раздражении заднего гипоталамуса. Это позволяет предположить наличие реципрокного взаимоотношения между передней и задней областями гипоталамуса и его значение для регуляции чередования различных фаз цикла бодрствование—сон. По данным Т.Н. Ониани, в цикле бодрствование— сон мультинейронная активность ретикулярной формации (активирующей системы) и преоптической области гипоталамуса (тормозной системы) меняется реципрокно (рис. 34). Представление о существовании двух центров, регулирующих цикл бодрствование— сон, согласуется с данными В. Гесса. Он показал, что раздражение задней области гипоталамуса вызывает симптомы, характерные для активации симпатической Рис. 34. Реципрокные отношения мультинейронной активности ретикулярной формации и преоптической области гипоталамуса в различных фазах цикла «бодрствование— сон». Активность нейронов ретикулярной формации во время бодрствования (а) и во время медленно-волновой фазы сна (б), активность нейронов преоптической области во время медленного сна (в) и во время бодрствования (г); калибровка: 40 мкв, 50 мс (по Т.Н. Ониани, 1983)
нервной системы (эрготропная система). На раздражение зоны в преоптической области и переднем гипоталамусе возникают симптомы, характерные для активации парасимпатической нервной системы (трофотропная система). Сами зоны находятся в реципрокном взаимоотношении. Однако гипоталамус занимает лишь низший уровень в лимбической системе, обеспечивающей регуляцию мотивационного поведения. В частности, он находится, под контролем миндалины (амигдалы). Базолатеральная область миндалины тормозит пищевое поведение, по-видимому, через вентромедиальный гипоталамус («центр насыщения»). На это указывает увеличение активности нейронов этой области гипоталамуса при стимуляции латерального ядра амигдалы. Дорзомедиальная миндалина оказывает на пищевое поведение облегчающее влияние прямо через активацию латерального гипоталамуса либо через торможение вентромедиального гипоталамуса, в результате чего имеет место высвобождение латерального гипоталамуса от тормозящего влияния со стороны вентромедиального ядра. Функции, осуществляемые миндалиной, в большей мере зависят от факторов внешней среды и ситуации удовлетворения потребностей, чем функции гипоталамуса. Миндалина скорее важна для различения вкусной и невкусной, новой и знакомой пищи, т. е. для формирования ответов на экстероцептивные пищевые стимулы. Согласно П.В. Симонову, главная функция миндалины — выделение доминирующей потребности, подлежащей первоочередному удовлетворению. Экспериментальным подтверждением этого положения является нарушение после двустороннего разрушения миндалин, выработки у крыс условнорефлекторного переключения разнородных рефлексов, по Э. А. Асратяну (когда один и тот же сигнал утром подкрепляется пищей, а вечером — болевым раздражением). П.В. Симонов полагает, что выбор поведения осуществляется «взвешиванием» конкурирующих эмоций, порожденных конкурирующими потребностями. Гипоталамус имеет тесные связи с лобной корой, которую, по-видимому, можно рассматривать как продолжение лимбической системы на корковом уровне. Во фронтальной коре найдены нейроны, по своим функциям сходные с глюкозочувствительными и глюкозонечувствительными нейронами латерального гипоталамуса. Нейроны ростральной дорзолатеральной префронтальной коры у обезьян преимущественно участвуют в процессах мотивации, а нейроны каудальной части — в сенсорном анализе вкуса и качества пищевого подкрепления. Нейрохимия мотивации Объединение различных структур мозга в функциональную систему, обеспечивающую реализацию определенной мотивации, имеет свою нейрохимическую основу. На это указывают уже упомянутые данные П.К. Анохина о химической и ЭЭГ-специфичности мотивационного возбуждения в составе различных типов поведения. Биохимической основой формирования мотиваций различного биологического качества является несколько групп биологически активных веществ. Прежде всего это — нейромедиаторы (ацетилхолин, серотонин, дофамин, норадреналин и др.), которые выделяются нервными окончаниями и служат посредниками в процессе синаптической передачи. Другую группу составляют гормоны, секретируемые железами внутренней секреции, и нейрогормоны, вырабатываемые нейросекреторными клетками нервной ткани. Гормоны и нейрогормоны поступают в кровь, гемолимфу, в тканевую и спиномозговую жидкость и оказывают дистантное длительное регулирующее действие на ЦНС и висцеральные органы. К последним относятся катехоламины (ДА, НА), серотонин, а также вещества полипептидной природы (вазопресин, окситоцин и др.). Большое внимание исследователей привлекают так называемые нейропептиды — особый класс веществ, обладающих исключительно высокой биологической активностью. Нейропептиды — относительно простые по своему строению соединения, состоящие из нескольких остатков аминокислот (от двух до нескольких десятков), синтезируются мозговой тканью (но и не только ею). Их главная особенность — полифункциональность: каждый нейропептид имеет необычайно широкий спектр действий, одновременно участвуя в регуляции множества мозговых функций. Полифункциональность, многоадресность пептидов позволяют им оказывать регулирующее влияние на организм в целом, захватывая как центральные, так и периферические механизмы, обеспечивая тем самым организацию целостных поведенческих актов. Частично это их свойство обусловлено их способностью взаимодействовать на субклеточно-молекулярном уровне с «классическими» нейромедиаторами и информационными макромолекулами (ДНК, РНК и белками). Отсюда нейропептиды, по-видимому, могут быть универсальными механизмами интеграции, возникшими на самых ранних этапах эволюции. Пептиды очень быстро распадаются в тканях. Длительность их жизни после высвобождения из мест синтеза или хранения измеряется минутами. Однако они способны вызывать отставленные эффекты, многие из которых обнаруживаются спустя несколько суток. По-видимому, пептиды могут запускать цепь каких-то процессов, которые развертываются уже без их участия, т. е. в этих случаях пептиды выполняют лишь пусковую функцию. Роль нейропептида в интеграции нервных образований в функциональную систему пищевого поведения показана К.В. Судаковым. В его опытах кроликам вживляли канюлю в левый боковой желудочек мозга, биполярные электроды в латеральный отдел гипоталамуса (центр голода) слева и вентромедиальное ядро гипоталамуса (центр оборонительных реакций) справа. Электрораздражение латерального отдела гипоталамуса вызывало у этих животных активное потребление пищи, а стимуляция вентромедиального ядра — пассивно-оборонительное поведение. Введение через канюлю в боковой желудочек 8-азагуа-нина и митомицина С блокировало стадию транскрипции (считывания) белков с рибонуклеиновой кислоты, а введение циклогексимида и пурацимина нарушало процесс транскрипции РНК с ДНК. Прерывание синтеза белков в клетке на любой его стадии сопровождалось избирательным разрушением пищедобывательного поведения, вызываемого как естественным путем через пищевую депривацию, так и электрической стимуляцией «центра голода» в латеральном гипоталамусе. При этом не страдали оборонительные реакции кролика, вызываемые электрическим раздражением вентромедиального гипоталамуса. Через некоторое время происходило полное восстановление пищевых реакций на стимуляцию гипоталамуса, а также естественного пищевого поведения. Длительность периода восстановления (от 1 до 200 часов) зависела от дозы введенного блокатора. Далее животным, у которых отмечалось полное угнетение пищевого поведения на фоне блокады синтеза белков, дополнительно вводили различные нейропептиды (АКТГ; пептид, вызывающий дельта-сон; октапептид холециетокина и пентогастрин). Оказалось, что только инъекция пентогастрина в дозе 1-2 мг/кг восстанавливает как естественное пищевое поведение, так и пищевые реакции, вызываемые электрической стимуляцией «центра голода». Сам пентогастрин блокируется антигастриновыми иммуноглобулинами (рис. 35). Исследователи полагают, что раздражение «центра голода» латерального отдела гипоталамуса и естественное пищевое возбуждение, по-видимому, активируют аппарат генетической памяти различных отделов ЦНС, в результате чего из клеток этих структур в межклеточное пространство выделяется пептидный фактор, имеющий структуру, близкую к гастрину. И этот нейропептид, по-видимому, играет роль «тригера» для функциональной системы питания. Рис. 35. Регуляция пищевого мотивационного возбуждения на молекулярном уровне. Возбуждение гипоталамуса с возникновением пищевой потребности активирует генный аппарат нейронов. В результате этого выделенный в межклеточную среду нейропептид (пентогастрин) вызывает пищедобывательное поведение (по К. В. Судакову, 1986)
Особая роль пентогастрина в инициации пищевого поведения показана и другими исследователями. Пентогастрин вызывает у накормленных кроликов дополнительный прием пищи. При этом нейроны латерального гипоталамуса начинают разряжаться ритмически с периодом 100-200 мс, т. е. их электрическая активность приобретает тип, характерный для состояния голода. Высокая концентрация вещества пептидной природы обнаружена у человека и животных в области гипоталамуса и в структурах лимбической системы, что тоже указывает на участие нейропептидов в центральных механизмах биологических мотиваций. Показано тормозное влияние ангиотензина II и пептида, вызывающего в ЭЭГ медленноволновый сон (дельта-сон), на оборонительные реакции, вызываемые электрической стимуляцией гипоталамуса, и на естественное оборонительное поведение кролика. Внутривенное введение нейропептида — субстанции «Р» — на 35% повышает порог вызова оборонительных реакций при стимуляции вентромедиального ядра гипоталамуса. Ее внутрибрюшинное введение значительно уменьшает число внезапных смертей у крыс в условиях эмоционального стресса, порождаемого иммобилизацией животного. Пищевая и питьевая мотивация возрастает под влиянием различных опиатов (нейропептидов), к которым относятся морфин и др. Антагонисты опиатов (налаксон, каптрексон) оказывают противоположный эффект — снижают потребление воды и пищи. Эмоции. Движение. В структуре мотивации особую функцию выполняет ее эмоциональный компонент. Эмоция, возникающая в составе мотивации, играет важную роль в определении направленности поведения и способов его реализации. Эмоция — особая форма психического отражения, которая в форме непосредственного переживания отражает не объективные явления, а субъективное к ним отношение. Особенность эмоций состоит в том, что они непосредственно отражают значимость действующих на индивид объектов и ситуаций, обусловленную отношением их объективных свойств к потребностям субъекта. Эмоции выполняют функции связи между действительностью и потребностями. Эмоции охватывают широкий круг явлений. Так, П. Милнер считает, что хотя и принято отличать эмоции (гнев, страх, радость и т. п.) от так называемых общих ощущений (голода, жажды и т. д.), тем не менее они обнаруживают много общего и их разделение достаточно условно. Одной из причин их различения является разная степень связи субъективных переживаний с возбуждением рецепторов. Так, переживание жары, боли субъективно связывается с возбуждением определенных рецепторов (температурных, болевых). На этом основании подобные состояния обычно и обозначаются как ощущения. Состояние же страха, гнева трудно связать с возбуждением рецепторов, и поэтому они обозначаются как эмоции. Другая причина, по которой эмоции противопоставляются общим ощущениям, состоит в нерегулярном их возникновении. Эмоции часто возникают спонтанно и зависят от случайных внешних факторов, тогда как голод, жажда, половое влечение возникают с определенными интервалами. Однако и эмоции, и общие ощущения возникают в составе мотивации как отражение определенного состояния внутренней среды, через возбуждение соответствующих рецепторов. Поэтому их различие условно и определяется особенностями изменения внутренней среды. Вместе с тем существует и иная точка зрения. Так, П. Фресс считает, что, хотя и существует единый континуум внутренних переживаний — от слабых чувств к сильным, только сильные переживания могут быть названы эмоциями. Их отличительной чертой является дезорганизующее влияние на текущую деятельность. Именно эти сильные чувства и обозначаются как эмоции. Эмоции развиваются, когда мотивация становится слишком сильной по сравнению с реальными возможностями субъекта. Их появление ведет к снижению уровня адаптации. Согласно этой точке зрения эмоции — это страх, гнев, горе, иногда радость, особенно чрезмерная радость. А удовольствие уже не является эмоцией. Хотя, как в случае сексуального удовольствия, оно может сопровождаться весьма интенсивными вегетативно-соматическими реакциями. Так, радость может стать эмоцией, когда из-за ее интенсивности мы теряем контроль над собственными реакциями: свидетельством тому являются возбуждение, бессвязная речь и даже безудержный смех. Такое сужение понятия эмоции соответствует представлению, выраженному в активационной теории Д. Линдсли, согласно которой эмоции соответствуют локальному участку на верху шкалы активации с наиболее высоким ее уровнем. Их появление сопровождается ухудшением выполняемой деятельности. Не все субъективные переживания относятся к эмоциям и по классификации эмоциональных явлений А.Н. Леонтьева, Он различает три вида эмоциональных процессов: аффекты, собственно эмоции и чувства. Аффекты — это сильные и относительно кратковременные эмоциональные переживания, сопровождающиеся резко выраженными двигательными и висцеральными проявлениями. У человека аффекты вызываются как биологически значимыми факторами, затрагивающими его физическое существование, так и социальными, например, социальными оценками, санкциями. Отличительной особенностью аффектов является то, что они возникают в ответ на уже фактически наступившую ситуацию. В отличие от аффектов собственно эмоции представляют собой более длительное состояние, иногда лишь слабо проявляющееся во внешнем поведении. Они выражают оценочное личностное отношение к складывающейся или возможной ситуации. Поэтому они способны, в отличие от аффектов, предвосхищать ситуации и события, которые реально еще не наступили. Они возникают на основе представлений о пережитых или воображаемых ситуациях. Третий вид эмоциональных процессов — это так называемые предметные чувства. Они возникают как специфическое обобщение эмоций и связаны с представлением или идеей о некотором объекте — конкретном или отвлеченном (например, чувство любви к человеку, к родине, чувство ненависти к врагу и т. д.). Предметные чувства выражают устойчивые эмоциональные отношения. Таким образом, наименее ясным остается вопрос о взаимоотношении эмоций как более узкого класса явлений, характеризующихся яркостью субъективных переживаний, с теми переживаниями, эмоциональная насыщенность которых менее выражена. Последние характерны для очень широк
|
||||
|
|
Последнее изменение этой страницы: 2016-12-15; просмотров: 491; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.89.181 (0.013 с.) |