Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Принцип конкурентного исключенияСодержание книги
Поиск на нашем сайте
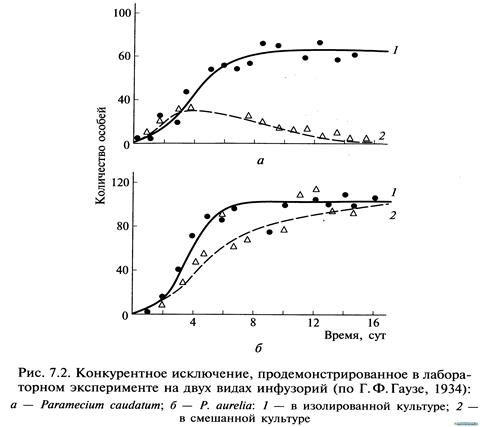
Внутривидовая конкуренция сильнее межвидовой, но правило конкуренции распространяется и на последнюю: конкуренция между двумя видами тем сильнее, чем ближе их потребности. В крайнем случае можно допустить, что два вида с совершенно одинаковыми потребностями не могут существовать вместе: один из них через какое-то время обязательно будет вытеснен. Это положение получило статус закона, известного как принцип конкурентного вытеснения (исключения), или принцип Гаузе — в честь Г. Ф. Гаузе, впервые продемонстрировавшего это явление в экспериментах с инфузориями (рис. 7.2). При изолированном содержании численность популяций обоих видов инфузорий увеличивается до тех пор, пока не достигнет максимума. В смешанной культуре популяция второго вида (рис. 7.2, б, кривая 2) растет быстрее, чем первого (рис. 7.2, а, кривая 2), что приводит к ограничению ее численности. Если не произойдет изменений в параметрах конкуренции, то конкурентное исключение (вымирание первой популяции) — лишь вопросвремени.
50.Понятия экосистемы и биогеоценоза, структурно-функциональная организация биогеоценоза.
Любая совокупность организмов, существующих в определенном объеме пространства, и их связей между собой и окружающей средой определяется как экологическая система, или экосистема. Термин «экосистема» впервые предложил английский эколог А. Тэнсли в 1935 г., хотя представления об экосистемах как особой форме взаимоотношений организмов со средой своего обитания возникли значительно раньше. Легко видеть, что понятие экосистемы является очень широким и не имеет ранга размерности. Экосистемой можно назвать любой объем пространства, в котором существуют живые организмы, хотя бы один. Ей может быть и капля воды с одной амебой, пруд, лесной массив, пилотируемый космический корабль и вся Биосфера Земли. В 1944 г. русский геоботаник В. Н. Сукачев предложил более конкретный термин «биогеоценоз». Он определяется как участок земной поверхности (в том числе и водной) с определенной совокупностью живых организмов и вполне однородными условиями среды, достаточно отделенный от других участков, в пределах которого осуществляется более-менее замкнутый процесс круговорота веществ. Примерами биогеоценозов являются пруд, луг, лесной массив, болото и т. д. Термины «экосистема» и «биогеоценоз» достаточно близки, но не полностью тождественны, поскольку понятие биогеоценоза значительно более конкретно, чем экосистемы. Например, капля воды и Биосфера являются экосистемами, но не биогеоценозами. Однако поскольку термин «экосистема» легче произносить, в дальнейшем мы будет употреблять его, но понимать в смысле биогеоценоза. Существование всех экологических систем обусловлено постоянным потоком энергии и круговоротом вещества в них. Все организмы на основании своей функциональной роли в экосистемеподразделяются на три важнейшие группы – продуценты, консументы и редуценты. Продуценты, или автотрофные организмы, создают органические вещества из неорганических в процессах фотосинтеза и хемосинтеза. Часть электромагнитной энергии солнечного излучения при этом переходит в энергию химических связей синтезируемых органических веществ. Консументы, или гетеротрофные организмы, не способны создавать самостоятельно органические вещества из неорганических, поэтому они используют для своей жизнедеятельности органические вещества, созданные автотрофными организмами.
Редуцентами являются преимущественно одноклеточные гетеротрофные организмы (бактерии, протисты), а также некоторые многоклеточные (грибы, почвенные черви и др.), которые разлагают мельчайшие органические остатки до простейших неорганических соединений – H2О, CO2, N2 и др., вновь возвращая их в экосистему. Выделяющийся при фотосинтезе кислород используется для дыхания самих растений, а также гетеротрофных организмов. Тепловая энергия, выделяющаяся при дыхании автотрофных и гетеротрофных организмов, рассеивается в пространстве, повышая энтропию окружающей среды. Образующиеся в процессах жизнедеятельности гетеротрофных организмов углекислый газ и вода используются автотрофными организмами в процессе фотосинтеза. Впервые целостные представления о закономерностях трансформации вещества энергии в экологических системах разработал шведский ученый Р. Линдеман, который в 1942 г. опубликовал работу о превращениях солнечной энергии в одном из арктических озер Швеции. Тем самым были заложены основы нового направления экологии – экологической биоэнергетики, которая оказала значительное воздействие на прогресс экологии и биологии в целом. В дальнейшем это направление интенсивно разрабатывалось многими учеными во всем мире. Среди них необходимо особо отметить Г. Г. Винберга, который в 1950–1960 гг. работал в Беларуси и стал основоположником белорусской экологической школы. Продуценты, или автотрофные организмы, являются основой существования всех экологических систем и биосферы Земли в целом, поскольку только они способны создавать органические вещества из неорганических впроцессах фотосинтеза и хемосинтеза. По этому признаку продуценты подразделяются на фото- и хемоавтотрофов.
Трофическая структура экосистем. Процессы трансформации вещества и энергии в экосистеме происходят преимущественно посредством трофических связей (от греческого трофос – пища), т. е. в результате поедания одних организмов другими. Совокупность трофических связей в экосистеме образует трофические, или пищевые, цепи. Совокупность организмов, занимающих одинаковое положение в трофических цепях относительно их начала, называется трофическим уровнем. Обычно каждый организм имеет несколько источников питания, находящихся на разных трофических уровнях, и сам может использоваться как объект питания организмами с различных уровней. Поэтому цепи питания в большинстве экосистем многократно разветвляются и образуют трофические сети. Различают пастбищные, детритные и паразитические пищевые цепи. Начальным звеном пастбищных цепей являются живые автотрофные (фотосинтезирующие или хемосинтезирующие) организмы. В пастбищных пищевых цепях первый трофический уровень образуют продуценты – автотрофные организмы. Последующие трофические уровни образуют гетеротрофные организмы. Второй трофический уровень составляют растительноядные организмы (консументы первого порядка) – животные, гетеротрофные протисты и бактерии. Третий и последующие трофические уровни (консументы второго, третьего и т.д. порядка) образуют хищные организмы – почти исключительно животные. Обычно разные трофические уровни в экосистемах не разделены в
пространстве. Однако в некоторых случаях они достаточно четко дифференцированы. Например, это имеет место в геотермальных источниках суши, в которых температура воды у изливов горячих подземных вод достигает 50–60 оС и более. Автотрофные организмы, преимущественно цианобактерии, образуют здесь специфические перифитонные сообщества («маты»), которые распространены при температурах свыше 40–45 оС, поскольку при более низких температурах они не выживают. С другой стороны, гетеротрофные организмы (моллюски, членистоногие и др.) в геотермальных источниках не встречаются при температурах более 33–36 оС, поэтому они питаются фрагментами мата, сносимого течением воды в зоны с более низкой температурой. Отсюда в таких геотермальных источниках четко выделяется автотрофная зона, где распространены только автотрофные организмы, и гетеротрофная зона, где автотрофные организмы отсутствуют и встречаются только гетеротрофные организмы (Хмелева, Голубев, Лаенко, 1984). В океанах и морях автотрофные организмы росли) существуют только до глубины проникновения света (максимум до 150–200 м). Гетеротрофные организмы, обитающие в более глубоких слоях воды, ночью поднимаются к поверхности, чтобы питаться водорослями, а утром вновь уходят на глубину, совершая суточные вертикальные миграции протяженностью до 500–1000 м. В свою очередь, с наступлением утра гетеротрофные организмы из еще более глубоких слоев поднимаются наверх, чтобы питаться организмами, опускающимися из вышележащих слоев. Таким образом, в океанах и глубоких морях существует своеобразная «пищевая лестница», благодаря которой органическое вещество, создаваемое автотрофными организмами в поверхностных слоях воды, переносится по цепочке живых организмов до самого дна. Вторым важнейшим источником органических веществ для видов из глубоководных экосистем являются организмы, отмирающие в верхних слоях и опускающиеся на дно («дождь трупов»). Общая масса отмирающих организмов в Мировом океане за год достигает не менее нескольких сотен миллионов тонн. В этой связи некоторые экологи рассматривают всю водную толщу отдельных регионов морей и океанов как единый биогеоценоз. Другие ученые полагают, что условия среды в поверхностных и придонных слоях воды настолько различны, что они являются разными биогеоценозами. В детритных пищевых цепях, или цепях разложения, продуценты (т. е. автотрофные организмы) отсутствуют. Их началом является детрит, или отмершее органическое вещество, представляющее собой остатки отмерших живых организмов. Детрит образует первый трофический уровень детритной пищевой цепи. Пастбищные пищевые цепи распространены преимущественно с сухопутных и морских экосистемах. Детритные цепи преобладают в
континентальных водоемах. Значительная часть пищевых ресурсов в них создается не в результате фотосинтеза (автохтонное органическое вещество), а поступает туда из наземных экосистем (аллохтонное органическое вещество), например, в виде листового опада. Гетеротрофные организмы, питающиеся непосредственно детритом, называются детритофагами. В наземных экосистемах ими являются многие виды насекомых, червей и др. Крупные детритофаги, к которым относятся некоторые виды птиц (грифы, вороны и т. д.), млекопитающих (гиены и прочие), а также многочисленные виды насекомых называют падальщиками. В водных экосистемах наиболее распространенными детритофагами являются членистоногие – водные насекомые и их личинки и ракообразные. Детритофагами питаются другие, более крупные гетеротрофные организмы, которые, в свою очередь, могут служить пищей для хищников. Детритные пищевые цепи имеют огромное значение в природе. Если бы их не было, то вся поверхность Земли быстро бы покрылась многокилометровым слоем отмерших организмов. Большое значение в разложении отмерших остатков растительного происхождения, особенно древесины, имеют грибы. Мельчайшие частицы отмершего органического вещества, которые не в состоянии усвоить детритофаги, разлагают бактерии и протисты, обладающие благодаря своим мельчайшим размерам очень высокой интенсивностью метаболизма. По этой причине они разлагают органические остатки до простых неорганических соединений (углекислый газ, вода, аммиак, азот и др.). Эти вещества могут возвращаться в экосистему и вновь принимать участие в процессах круговорота веществ. Как и в пастбищных цепях, последний трофических уровень детритных пищевых цепей образуют редуценты- микроконсументы. Практически любой вид живых организмов имеет своих паразитов. Начальным звеном паразитических пищевых цепей является свободноживущий организм-хозяин. На нем паразитируют паразиты первого порядка, а на них в, свою очередь, паразиты второго порядка и т. д. Аскариды, паразитирующие на пойкилотермных организмах, являются паразитами первого порядка, паразитирующие в клетках аскарид одноклеточные эукариоты –паразитами второго порядка, обитающие в них бактерии – паразитами третьего порядка, паразитирующие в бактериях вирусы (бактериофаги) – четвертого порядка и т. д. В экологических системах все перечисленные типы пищевых цепей не существуют изолированно, а тесно взаимосвязаны. Так, отмирающие организмы из пастбищных пищевых цепей и неусвоенные остатки пищи живых организмов поступают в детритные цепи, поэтому в экосистемах формируется Y-образная
50.Любая совокупность организмов, существующих в определенном объеме пространства, и их связей между собой и окружающей средой определяется как экологическая система, или экосистема. Термин«экосистема» впервые предложил английский эколог А. Тэнсли в 1935 г., хотяпредставления об экосистемах как особой форме взаимоотношений организмовсо средой своего обитания возникли значительно раньше.Легко видеть, что понятие экосистемы является очень широким и не имеетранга размерности. Экосистемой можно назвать любой объем пространства, вкотором существуют живые организмы, хотя бы один. Ей может быть и капляводы с одной амебой, пруд, лесной массив, пилотируемый космический корабльи вся Биосфера Земли.
В 1944 г. русский геоботаник В. Н. Сукачев предложил более конкретный термин «биогеоценоз». Он определяется как участок земной поверхности (в том числе и водной) с определенной совокупностью живых организмов и вполне однородными условиями среды, достаточно отделенный от других участков, в пределах которого осуществляется более-менее замкнутый процесс круговорота веществ. Примерами биогеоценозов являются пруд, луг, лесноймассив, болото и т. д.Термины «экосистема» и «биогеоценоз» достаточно близки, но неполностью тождественны, поскольку понятие биогеоценоза значительно болееконкретно, чем экосистемы. Например, капля воды и Биосфера являютсяэкосистемами, но не биогеоценозами. Однако поскольку термин «экосистема»легче произносить, в дальнейшем мы будет употреблять его, но понимать всмысле биогеоценоза. Существование всех экологических систем обусловлено постоянным потоком энергии и круговоротом вещества в них. Все организмы на основании своей функциональной роли в экосистеме подразделяются на три важнейшие группы – продуценты, консументы и редуценты. Продуценты, или автотрофные организмы, создают органические вещества из неорганических в процессах фотосинтеза и хемосинтеза. Часть электромагнитной энергии солнечного излучения при этом переходит в энергию химических связей синтезируемых органических веществ. Консументы, или гетеротрофные организмы, не способны создавать самостоятельно органические вещества из неорганических, поэтому они используют для своей жизнедеятельности органические вещества, созданные автотрофными организмами. Редуцентами являются преимущественно одноклеточные гетеротрофные организмы (бактерии, протисты), а также некоторые многоклеточные (грибы, почвенные черви и др.), которые разлагают мельчайшие органические остатки до простейших неорганических соединений – H2О, CO2, N2 и др., вновь возвращая их в экосистему.
51. Различные типы объединений живых организмов. Живые организмы в экосистеме группируются не хаотически, а объединяются в определенные группы по причине своих биологических особенностей и воздействия определенных факторов внешней среды. Выделяют несколько типов подобных объединений, отличающихся разной степенью интегрированности. 1. Общественные группировки, объединяющие особей одного вида. В их образовании важнейшая роль принадлежит не факторам внешней среды, а социальным отношениям и поведенческим реакциям. Они образуются у насекомых, птиц, млекопитающих. Их примерами являются пчелиный рой, муравейник, косяк рыб, прайд львов, сообщество обезьян и т. п. Общественные группировки относительно независимы от окружающей среды, они сохраняются даже тогда, когда условия среды меняются. Например,муравейник может сохраняться на протяжении нескольких десятков лет. 2. Скопления – собрание особей, относящихся к одному или нескольким видам. Они возникают исключительно под воздействием одного или нескольких внешних факторов, выступающих в качестве центра притяжения.Скопления имеют случайный и временный характер. Между организмами,образующими скопление, нет устойчивых связей; оно разрушается, как только исчезает центр притяжения. Примером скопления может быть ночной рой насекомых, образованный особями разных видов, вокруг электрической лампы.Если лампу выключить, рой исчезает. 3. Активные паразитарные сообщества возникают благодаря привлечению одного животного другим; оно происходит между разными видами, но имеет исключительно односторонний характер. Например, любой живой организм имеет многочисленных паразитов и комменсалов, которые могут принадлежать к нескольким десяткам (если не сотням) видов. При гибели хозяина эти сообщества, как правило, распадаются. 4. Псевдообщественные сообщества – скопления, возникшие под действием тигмотаксисов, т. е. стремления организмов к контакту с определенным субстратом. Например, в водоемах одни группы организмов обитают только на твердых, а другие – на мягких грунтах. 5. Биоценоз. Этот термин предложил немецкий зоолог Карл Мёбиус в 1877 г. в работе о сообществах животных и растений, приуроченных к устричным банкам. Сам Мёбиус дал следующее пространное определение биоценозу – объединение живых организмов, соответствующее по своему составу, числу видов и особей некоторым средним условиям среды; объединение, в котором организмы связаны взаимной зависимостью и сохраняются благодаря постоянному размножению в определенных местах.… Если бы одно из условий отклонилось на некоторое время от обычной средней величины, изменился бы весь биоценоз …. Биоценоз также претерпел бы изменения, если бы число особей данного вида увеличилось или уменьшилось благодаря деятельности человека, или же один вид полностью исчез из сообщества, или, наконец, в его состав вошел новый». Синонимами термина «биоценоз» являются такие понятия, как «ассоциация», «сообщество» и «биотическое сообщество». Часто под биоценозом понимается не совокупность организмов, а совокупность популяций.Термин «сообщество» можно также применить для обозначения любой группы взаимодействующих популяций. Например, можно рассматривать сообщества лужи, дубового листа и т. д. Однако мы будет понимать это пространство в смысле биоценоза.Совокупность популяций отдельных видов, населяющих биоценоз, носит название биоценотическое сообщество, или биоценоз. Природная среда, в которой существует биоценоз, называется биотоп. Поэтому биогеоценоз =биоценоз + биотоп. Виды, образующие биоценоз, взаимно связаны друг с другом. Это отличает биоценоз от скоплений и псевдообщественных сообществ, где подобных связей нет или они очень слабы. Взаимосвязь эта так сильна, что изменения, касающиеся только одного вида, могут отразиться на всем биоценозе и даже вызвать его распад. Особенно это касается видов-эдификаторов, или «строителей сообщества», которые в результате своей жизнедеятельности создают специфическую среду обитания для других видов сообщества. Примером вида-эдификатора может быть ель в сообществе старого и густого елового леса. Кроны еловых деревьев образуют сомкнутый полог, под которым создается специфический микроклимат, характеризующийся резким снижением освещенности, особым температурным режимом и повышенной влажностью воздуха. В таких условиях могут существовать только тенелюбивые и влаголюбивые виды живых организмов. Биоценозы подразделяются на несколько уровней. 1. Основные сообщества. В Биосфере Земли их всего три – наземные, пресноводные и морские. 2. Биомы, или формации, природные комплексы, природные зоны. Биом – это однородное сообщество, не зависящее от состава растительности и занимающее обширную территорию – от миллионов до десятков миллионов км2.Причинами, обусловливающими видовой состав биома являются особенности климата, геологические факторы (преграды и пр.), эволюция живых организмов.Примерами биомов являются тундра, тайга, саванна, коралловые рифы,открытый океан, шельфовые зоны и т. п. Биомы в равной степени являются объектами изучения экологии и биогеографии. 3. Ассоциации, выделяемые в пределах биома, представляют собой локальные, хорошо очерченные группировки видов. Их примерами являются различные типы леса – хвойный, дубовый и т. п. 4. Синузии, или микроассоциации – некоторые очень небольшие сообщества, четко ограниченные в пространстве и сохраняющие свою индивидуальность. Ими могут быть виды, поселяющиеся на трупе крупного животного, стволе мертвого дерева и т.п. Отдельные синузии существуют очень непродолжительное время, однако синузии, постоянно возникающие в пределах одной ассоциации, имеют сходный видовой состав. 10.6. Таксономический состав экологических систем. Все экологические системы существуют в пространстве и времени благодаря происходящим в них процессам трансформации вещества и энергии. В этих процессах принимают участие все имеющиеся в экосистемах живые организмы. Все они значительно различаются по строению и особенностям метаболизма, в результате чего выполняют разные функции в экосистеме.На основе ряда фундаментальных особенностей строения и метаболизма все виды живых организмов на Земле подразделяются на ряд высших таксономических групп, которые имеют ранг доменов (domain), или надцарств и царств. Прежние достаточно простые схемы, выделяющие четыре или пять царств, в настоящее время кардинально пересмотрены. Сейчас все живые организмы подразделяются на три домена – Архебактери (Archaea), Бактерии (Bacteria) и Эукариоты (Eucaria). Каждый домен включает не менее пяти групп,ранг которых соответствует царству. 1. Архебактерии (несколько сотен видов) – аэробные и анаэробные прокариотные организмы. Обитают в глубовоководных районах Мирового океана близи выходов гидротермальных вод. Отдельные виды обитают при температуре свыше 114 оС. Способны к хемосинтезу. 2. Бактерии (около 80 тысяч видов) – прокариотные организмы, в клетках которых отсутствуют ядра, митохондрии и хлоропласты. К ним относятся ряд групп бактерий и цианобактерии. Большинство – гетеротрофы с сапрофитнымспособом питания. Многие виды способны к фотосинтезу или хемосинтезу. Неменее 2–3 тысяч видов являются облигатными анаэробами. 3. Эукариоты (более 2 миллионов видов) – одноклеточные и многоклеточные организмы, в клетках которых имеются ядра и митохондрии, а у автотрофных форм – и хлоропласты. К ним относится ряд царств одноклеточных, которых ранее объединяли в царство Протистов – Жгутиковые, Ресничные и Микроспоридии, а также «коронные» царства, включающие многоклеточные организмы, – Грибы, Растения и Животные. Все животные, грибы и многие протисты являются гетеротрофами. Способы их питания различаются от сапрофитного – у грибов и животных-эндопаразитов до голозойного (т. е. питания оформленной пищей) – у свободноживущих и протистов. Абсолютное большинство растений и многие протисты являются автотрофами. Топические группировка биогеоценозов. В наземных экосистемах растительные сообщества автотрофных организмов обычно представлены сосудистыми растениями – покрытосемянными, голосемянными, папоротникообразные, хвощами, плаунами, мохообразными, лишайниками. Эти сообщества обычно подразделяются на пять вертикальных ярусов. Первый, самый верхний, ярус составляют высокие деревья, кроны которых часто смыкаются, образуя сплошной полог. Второй ярус образуют кустарники(крушина, бузина, малина и др.), третий – кустарнички (черника, брусника,голубика, клюква, вереск и пр.). Четвертый ярус формируют травянистые растения и, наконец, пятый ярус – напочвенные мхи и лишайники.В некоторых сообществах отдельные ярусы могут отсутствовать. Например, в старых и густых еловых лесах часто имеется лишь первый ярус. Сомкнутые кроны елей создают хорошее затенение, что затрудняет развитие не только кустарниковой растительности, но и собственного подроста, а мощный слой хвойного опада препятствует формированию остальных ярусов.В высоких и густых тропических лесах микроклимат отдельных ярусов может существенно различаться. В приземном слое здесь очень высокая влажность и низкая освещенность, а температура в течение суток практически не изменяется. Напротив, микроклимат у верхушек деревьев, характеризующийся очень высокой освещенностью, более низкой влажностью и значительными суточными колебаниями температуры, больше напоминает климат саванны.В каждом из ярусов можно найти своеобразные группировки гетеротрофных организмов, преимущественно насекомых, но иногда к ним приурочены определенные виды птиц, млекопитающих (например, белки,ленивцы) и даже амфибий (квакши) и рептилий (летучий дракон). В водных экосистемах существуют специфические биоценозы донных организмов и обитающих в водной толще, называемые соответственно бентос и планктон. Далее сообщество донных бактерий называется бактериобентос,растений – фитобентос и животных – зообентос. По тому же принципу выделяют бактериопланктон, фитопланктон и зоопланктон. В морских экосистемах фитобентос образуют исключительно многоклеточные водоросли – красные, бурые и др.; виды цветковых растений,например, зостера Zostera marina, здесь единичны. В фитобентосе пресных водоемов, напротив, доминируют погруженные и полупогруженные цветковые растения, а многоклеточные водоросли (харовые, зеленые и др.) имеют второстепенное значение. Фитопланктон в обоих случаях представлен одноклеточными водорослями; в морских экосистемах доминируют диатомовые, а в пресноводных – зеленые водоросли. К пресноводному фитопланктону (а не к бактериопланктону) также обычно относят и цианобактерий. Последние периодически, обычно во второй половине лета, могут достигать очень высокой численности и биомассы («цветение воды») и тогда вытесняют остальные виды водорослей. К зоопланктону относят достаточно мелких животных, не способных противостоять течению воды, хотя многие их виды в стоячей воде способны к активному плаванию. В зоопланктоне пресных водоемов представлены преимущественно мелкие формы – веслоногие и ветвистоусые ракообразные и коловратки. Зоопланктон морских водоемов значительно более разнообразен. В нем помимо мелких ракообразных (преимущественно веслоногих, мизид и эуфаузиид), аппендикулярий и других, присутствуют и значительно более крупные медузы и сальпы; последние, наряду с аппендикуляриями, относятся к разным подтипам хордовых. Значительную долю в зоопланктоне континентальных и морских водоемов составляют личинки многочисленных видов бентосных животных, выполняющие функцию расселения, а также личинки и мальки рыб. Зообентос континентальных водоемов в видовом отношении достаточно однообразен – в нем доминируют брюхоногие и двустворчатые моллюски, членистоногие (почти исключительно ракообразные и насекомые и их личинки) и малощетинковые черви. Напротив, в зообентосе морских водоемов представлено большинство таксонов водных животных, поэтому его можно здесь охарактеризовать лишь в самых общих чертах. В первую очередь необходимо отметить коралловых полипов, которые как виды-эдификаторы в экосистемах коралловых рифов создают среду обитания для многочисленных видов морских водорослей, беспозвоночных и рыб. Значительная роль в морском зообентосе также принадлежит двустворчатым и брюхоногим моллюскам, кольчатым червям, ракообразным, иглокожим, асцидиям. В зообентосе глубоководных океанических экосистем доминируют погонофоры.Сообщество водных организмов, постоянно обитающих в толще воды, но способных активно плавать и противостоять течению, называется нектон. Нектон пресных водоемов образуют почти исключительно рыбы. В нектонеморей и океанов наряду с ними присутствуют головоногие моллюски (кальмары,осьминоги) и морские млекопитающие (киты, дельфины, дюгони). Сообщества, образующие обрастания на вертикальных естественных (скалы, камни, подводные растения) и искусственных (сваи, подводные сооружения, днища судов) подводных субстратах, называются перифитоном. Он подразделяется на зоо- и фитоперифитон. Сообщества зоо- и фитоперифитона континентальных и морских водоемов в таксономическом отношении достаточно сходны. В зооперифитоне присутствуют губки, мшанки, моллюски, нематоды, полихеты и др. Для морских сообществ характерно также наличие усоногих ракообразных и иглокожих. Видами-эдификаторами в разных сообществах зооперифитона могут являться двустворчатые моллюски, образующие устричные и мидиевые банки,губки, мшанки и др. Фитоперифитон образуют различные группы многоклеточных водорослей.Сообщества перифитона могут приносить значительный вред человеку.В них имеется ряд видов животных, которые разрушают не только подводные деревянные сооружении (например, двустворчатый моллюск – корабельный червь), но даже металлические конструкции – некоторые виды морских ежей.Обрастания на днищах кораблей значительно снижают их скорость.
52. Свойство экосистемы создавать в процессе своего функционирования органическое вещество определяется как биологическая продуктивность. Органическое вещество, создаваемое в экосистеме, в результате прироста массы тела населяющих ее организмов, называется продукцией. Различают продукцию популяций отдельных видов, видовых сообществ, целых трофических уровней и всей экосистемы. Все органическое вещество, создаваемое автотрофными организмами в процессах фотосинтеза или хемосинтеза, называется валовой первичной продукцией (ВПП). Однако значительная часть синтезированного органического вещества сразу же расходуется ими в процессах метаболизма. Разность между энергией ВПП и расходом ее в метаболических процессах (М)является чистой первичной продукцией (ЧПП). Таким образом, ЧПП = ВПП – М. Чистая первичная продукция является тем самым органическимвеществом, которое накапливается в растениях. Из него в результате сложныхпроцессов, происходящих в растениях, образуются все их органы – стебли,листья, корни, цветки плоды и другие, которые могут использоваться в пищугетеротрофными организмами. Органическое вещество, создаваемое гетеротрофными организмами, находящимися на втором и последующих трофических уровнях, называется вторичной продукцией. Она равна сумме приростов массы их тела гетеротрофных организмов за определенный период времени. В большинстве случаев вторичная продукция, которую часто называют просто продукцией, рассчитывается для популяций отдельных видов. Этим она отличается от первичной продукции, которую возможно рассчитать для всего сообщества автотрофных организмов, составляющих данную экосистему. Количество органического вещества, покидающего тем или иным способом экосистему (сбор урожая, вылов рыбы, вылет наземных насекомых из их водных личинок и т. д.), называется конечной продукцией. Способы выражения биомассы и продукции. Общее количество живого вещества в экосистеме называется биомассой. Биомасса экосистемы образована всеми населяющими ее живыми организмами. Продукция экосистемы равна приросту ее биомассы за определенный промежуток времени. Биомасса и продукция экосистем могут быть выражены в различных единицах, однако предпочтительнее их выражать в одинаковых единицах.Биомасса экосистемы рассчитывается как произведение численности (или плотности) особей в экосистеме на их среднюю массу. Поскольку особи в экосистеме объединены в популяции отдельных видов, биомасса экосистемы равна сумме биомасс всех составляющих ее популяций. В свою очередь биомасса популяции равна сумме масс тела всех ее особей. Биомасса экосистемы является статическим показателем, который не имеет размерности времени. Напротив, продукция – динамический показатель,который определяется для какого-либо промежутка времени – суток, месяца,года и т. д. В наземных экосистемах биомасса и продукция обычно пересчитываются на единицу площади (г · м-2; т · км-2 · год-1 и т. д.). Одним из важнейших показателей, характеризующих продукционные возможности организмов, является Р/B -коэффициент, или отношение продукции (Р) какой-либо популяции или даже всего трофического уровня к биомассе (В) этой популяции или трофического уровня. Значения Р/B рассчитываются для определенного периода времени, обычно за сутки, месяц,сезон вегетации, год и т.д., поэтому имеют размерность соответственно год-1,сезон вегетации-1, месяц-1, сутки-1 и т. д. Суточный Р/B -коэффициент популяций гетеротрофных организмов часто называют удельной продукцией. В качестве биомассы при расчетах Р/B используют ее минимальное или, максимальное, но чаще среднее значение за тот период времени, для которого производится расчет.
53. Различают валовую П. п., равную общему кол-ву продуктов фотосинтеза за определённый отрезок времени, и чистую П. п., равную разности между валовой П. п. и той её частью, к-рая была затрачена в процессе дыхания.Все органическое вещество, создаваемое автотрофными организмами в процессах фотосинтеза или хемосинтеза, называется валовой первичной продукцией (ВПП). Методы определения первичной продукции. Чистая первичная продукция наземных экосистем обычно оценивают по годовому приросту биомассы растений. Чтобы определить ее величину за сезон вегетации, в конце этого сезона все растения, находящиеся на определенных территориях известной площади, удаляют и взвешивают. Поскольку обычно корни и подземные части растений не учитываются, определяют только массу их наземных органов – или чистую годовую наземную продуктивность. По отношению зерновым сельскохозяйственным культурам она будет равной суммарной массе собранного зерна, с
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-12-12; просмотров: 878; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.117.19 (0.021 с.) |