Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Химическое строение нуклеиновых кислот.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Нуклеиновые кислоты – это биологические полимеры, мономерами которых являются нуклеотиды. Каждый нуклеотид содержит азотистое основание определенного типа, пентозу (рибозу или дезоксирибозу) и фосфорную кислоту. Азотистые основания. Выделяют 2 вида азотистых оснований – пурины и пиримидины. Пурины содержат два кольца - 5-ти и 6-тичленное – это аденин и гуанин (рис.3).
Аденин Гуанин Рис. 3. Строение пуриновых азотистых оснований.
Пиримидины образованы одним 6-тичленным кольцом – это тимин, урацил и цитозин (рис.4).
Цитозин Тимин Урацил Рис. 4. Строение пиримидиновых азотистых оснований.
Основания принято обозначать первой буквой их названия: Аденин-А, Тимин-Т, Гуанин-Г, Цитозин-Ц, Урацил-У. Сахар – рибоза или дезоксирибоза (рис.5). В зависимости от сахара различают 2 вида нуклеиновых кислот – ДНК и РНК.
Рибоза Дезоксирибоза Рис. 5. Пентозы нуклеиновых кислот. Фосфорная кислота – придает нуклеиновым кислотам кислотные свойства (рис.6).
Рис. 6. Фосфорная кислота.
Рис. 7. Строение нуклеозида и нуклеотида.
Молекула нуклеотида ассиметрична. Начинается нуклеотид фосфорной кислотой, которая присоединяется к 5' – углероду собственного сахара, а к 1' – углероду сахара присоединяется азотистое основание (рис.7). Таким образом, в химическом строение нуклеотида заложено направление транскрипции, трансляции, репликации ДНК от 5 ' конца к 3 ' концу. ДНК и РНК всех живых организмов образованы сочетанием 4-х типов нуклеотидов (табл.1).
Таблица 1. Виды нуклеотидов ДНК и РНК.
Нуклеотиды соединяются в полинуклеотидную цепь прочными ковалентными фосфодиэфирными связями: к 3' углероду сахара предыдущего нуклеотида присоединяется остаток фосфорной кислоты следующего нуклеотида (рис.8).
Таким образом, полинуклеотидная цепь представляет из себя сахаро – фосфатный остов, к которому перпендикулярно присоединены азотистые основания. Дезоксирибонуклеиновая кислота. Строение ДНК. Схема строения ДНК была предложена в 1953г. Джеймсом Уотсоном и Френсисом Криком. Они избрали удачный подход, который обеспечил им успешное решение проблемы. Дж. Уотсон и Ф. Крик решили построить трехмерную модель ДНК, которая бы объясняла ее свойства и функции. За эту работу они получили Нобелевскую премию. Основные научные факты, которые использовали в своей работе Дж. Уотсон и Ф. Крик: 1. 1950 г. – английский биофизик Морис Уилкинс и его ученица Розалинда Франклин на рентгенограмме кристаллических волокон ДНК (рис.9) получили четкое подтверждение двойной спирали (крестообразный рисунок). Нуклеотиды соединяются с помощью фосфодиэфирной связи (5'→3'). (М. Сингер, П. Берг, 1998, с. 40)
Рис. 9. Рентгенограмма кристаллических волокон ДНК. («От молекул до человека», 1973, с. 91)
Кроме того, эти данные показали: a) Спираль имеет диметр 2 нм; b) Постоянный диаметр спирали предпологал, что в каждой цепи азотистые основания направлены внутрь спирали;
2. 1950 г. – английская группа А. Тоддa установила точную структуру связи между нуклеотидами в полинуклеотидной цепи – фосфодиэфирную связь (рис.8) 3. 1950-51 гг. – Э. Чаргафф проанализировал количественный состав ДНК и сформулировал несколько правил: a) Состав ДНК различных клеток, составляющий ткани и органы одного организма, всегда одинаков. b) Состав ДНК клеток организма с возрастом не изменяется. c) Состав ДНК клеток разных видов различен. d) Количество аденина равно количеству тимина, а количество гуанина равно количеству цитозина. e) Сумма пуриновых оснований равна сумме пиримидиновых: А+Г = Ц+Т. Эти закономерности, свидетельствовали о каких – то строгих принципах построения ДНК. 4. Кислотно-щелочное титрование ДНК показало, что ее структура стабилизируется водородными связями. Проанализировав огромное количество научных фактов Дж. Уотсон и Ф. Крик предложили модель строения ДНК. Они показали, что ДНК образована двумя вправозакрученными полинуклеотидными цепями. (рис. 10)
Различают три уровня организации ДНК. Первичная структура – одиночная полинуклеотидная цепь (рис.8). Вторичная структура – двойная вправозакрученная спираль, т.е. две полинуклеотидные цепи закручены вправо вокруг общей воображаемой оси (тип винтовой лестницы) (рис. 10, 11). В основе построения вторичной структуры ДНК лежат два принципа: 1. Принцип комплементарности (комплемент – взаимодополнение). Азотистому основанию одной цепи строго соответствует определенное азотистое основание другой цепи: А-Т, Г-Ц. Принцип комплементарности позволяет: а) заполнить максимально водородные связи, которые, располагаясь на всем протяжении молекулы ДНК с одной стороны, придают ей большую устойчивость, с другой стороны – гибкость. Между А и Т образуются две водородные связи, между Г и Ц – три (рис.11). б) сохранить по всей длине молекулы ДНК одинаковое расстояние между цепями (1,1 нм), что стабилизирует в целом молекулу ДНК, т.е. придает ей устойчивость. (Два пурина занимали бы слишком много места, а два пиримидина слишком мало, чтобы заполнить промежуток между двумя цепями) в) дублировать наследственную информацию, что повышает стабильность ее хранения, т.к. информация записана на двух цепях одновременно. Физико-химической основой комплементарности являются следующие факторы: 1. Пространственная конфигурация азотистых оснований: форма А соответствует форме Т, а Г – Ц. 2. Водородные связи 3. «Стекинг» взаимодействия. Вдоль спирали основания уложены стопками друг на друга и стабилизация спиральной структуры дополнительно обеспечивается межплоскостными взаимодействиями между ароматическими кольцами соседних оснований. Эти специфические контакты получили название стекинг – взаимодействия. Стекинг – взаимодействия возникают в результате Ван-дер-Ваальсовых сил и гидрофобных взаимодействий между соседними азотистыми основаниями уложенными друг над другом. 2. Принцип антипараллельности – обеспечивает компактизацию молекулы ДНК, свободное пространство одной цепи заполняется молекулярными структурами другой цепи (рис.10,11). Полинуклеотидные цепи антипараллельны друг другу, т.е. направление цепей: 5' → 3' 3' ← 5' Таким образом, молекула ДНК имеет следующие особенности: 1.число полинуклеотидных цепей равно 2 2.цепи образуют спирали по 10 пар нуклеотидов в каждом витке 3.двойные цепи закручены каждая вправо вокруг другой и вместе вокруг общей оси 4.сахарофосфатный остов снаружи, внутри под прямым углом расположены основания с интервалом 0,34 нм 5.цепи удерживаются вместе водородными связями между основаниями и стекинг - взаимодействиями 6.полинуклеотидные цепи комплементарны друг другу 7.диаметр спирали 2 нм 8.в структуре ДНК заложена возможность так называемой конвариантной репликации. Этим термином советский генетик Н.В. Тимофеев – Ресовский назвал способность живых организмов воспроизводить себе подобных. Следует добавить, что Дж. Уотсон и Ф. Крик описали одну из возможных форм существования ДНК – В-форму. Позже были открыты альтернативные двуспиральные структуры ДНК, что свидетельствует о полиморфизме ДНК (А,В,Z,С,Д,Е – формы). Формы различаются количеством нуклеотидных пар в витке, углом наклона плоскости азотистых основании к оси и т.д.(рис.12).
Эти формы ДНК обнаружены при определенных условиях: влажности, разных последовательностях нуклеотидов в цепи, присутствии ионов и т. д. Третичная структура ДНК характеризуется спирализацией и супер(сверх)спирализацией. (рис.20).
Уровни компактизации ДНК. Длина ДНК диплоидного набора хромосом человека составляет примерно 174 см., средняя длина ДНК одной хромосомы – 5 см. В ядре длина одной хромосомы составляет 0,5 – 1 микрон. Такая упаковка двойной спирали ДНК объясняется ее дальнейшей последовательной компактизацией.
Рис. 12. A-, B-, C- и D-формы ДНК (А. С. Коничев, Г. А. Севастьянова, 2005, с. 90)
1. Нуклеосомный уровень. Нуклеосома - это ДНК - гистоновый комплекс, который выглядит как частица дисковидной формы диаметром 11 нм. Впервые нуклеосомы были описаны в 1974г. А. Олинс и Д. Олинс. Каждая нуклеосома состоит из белкового кора или октамера и 2 оборотов фрагмента двухцепочечной ДНК (рис.13).
Белковый кор (сердцевина) содержит набор из 4 пар гистоновых белковН2А, Н2В, Н3, Н4. Это самые консервативные белки в любом геноме. Они практически одинаковы у гороха и у человека. Нуклеосомы связываются участками ДНК (линкерная ДНК) свободными от контакта с белковым кором. Укладка линкерного участка ДНК (60-80 п.н.) и соединение нуклеосом друг с другом идут с помощью гистона Н1. Молекула этого белка имеет центральную (глобулярную) часть и вытянутые «плечи». Центральная часть прикрепляется к специфическому участку на поверхности кора, вытянутые «плечи» соединяют соседние нуклеосомы. При этом ДНК наматывается на соседние коры каждый paз в противоположном направлении (рис. 14). Выделить нуклеосомы можно непродолжительной обработкой хромосом ферментами дезоксирибонуклеазами. При этом расщепляются участки состыковки нуклеосом. В геноме человека содержатся 1,5 х 107 нуклеосом. Нуклеосомный уровень повышает плотность упаковки ДНК в 7-10 раз. (Рис. 14, 20)
Рис.14. Модель нуклеосомной фибриллы. 2. Нуклеомерный уровень. Дальнейшая компактизация ДНК в составе хроматина связана с образованием нуклеосомных комплексов (рис. 15, 20).Образуется компактная хроматиновая фибрилла построенная либо по типу соленоида (спиральный тип укладки), либо по нуклеомерному типу (4-12 нуклеосом образуют глобулу).
Нуклеомерная укладка хроматина способствует укорочению нити ДНК примерно в 6 раз, а оба уровня приводят к компактизации ДНК в среднем в 50 раз (42-60). 3. Хромомерный уровень. Следующий этап компактизации ДНК связан с образованием петлеобразных структур, которые называются хромомерами (рис.16). При этом возможны два пути упаковки ДНК с помощью негистоновых белков:
Рис. 16. Хромомерный тип укладки хромосом.
Нить нуклеосом разбита на участки по 20 - 80 тысяч пар азотистых оснований (в среднем – 50 тысяч). В местах разбивки находятся молекулы – глобулы - негистоновых хромосомных белков. ДНК - связывающие белки узнают глобулы негистоновых белков и сближают их. Образуется устье петли. Средняя длина петли (300-400 нм) сходна у различных организмов (дрозофила и человек) и включает примерно 50 тысяч оснований. Такую петельную структуру называют интерфазной хромонемой. Хроматин типа «ламповых щеток» - это интерфазный эухроматин (рис.17.). Считают, что петли имеют связи с белками хромосомного каркаса, ядерного матрикса и белками ламины.
Рис. 17. Фрагменты хромосом типа «ламповых щеток» из ядра ооцита тритона. Можно видеть участки ДНК, образующие петли от центральной оси. (С. Гильберт, 1993, т. 2, с. 186)
Укорочение фибриллы на этом уровне происходит в среднем 25 раз, а на всех 3 уровнях в 1000-1500 раз.
4. Хромонемный уровень. При делении клеток идет дальнейшая компактизация хромосом - образование более крупных петель из хромомерной фибриллы. На поверхности упакованные молекулы ДНК несут множество белков, которые образуют подобие чехла. Если удалить этот чехол, то под электронным микроскопом можно отчетливо увидеть, что каждая хроматида построена из хроматиновых петель, отходящих от центральной оси. Диаметр такой упаковки 700 нм (рис. 18).
Рис.18. Хромонемный тип укладки хромосом.
5. Хромосомный уровень. Дальнейшая компактизация хромосом обеспечивается петельной укладкой хромонемной нити (рис.19.), что сокращает их длину примерно в 10 раз.
Рис.19. Хромосомный тип укладки.
Кроме того, компактизация хромосомы - важнейший процесс, связанный с точной передачей наследственной информации очередному поколению.
Свойства ДНК. Свойства ДНК определяются ее строением: 1. Универсальность - принципы построения ДНК для всех организмов одинаковы. 2. Специфичность - определяется соотношением азотистых оснований: А + Т, Г + Ц которое специфично для каждого вида. Так у человека оно составляет 1,35, у бактерий – 0,39 Специфичность зависит от: · количества нуклеотидов · вида нуклеотидов · расположение нуклеотидов в цепи ДНК
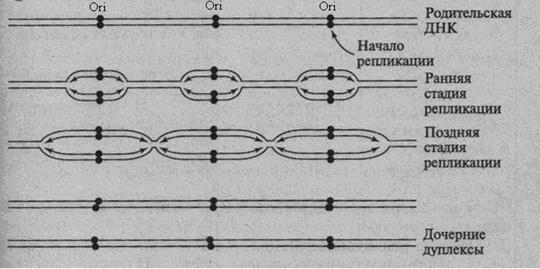
Единицей репликации является репликон. Репликон – это участок ДНК, где происходит репликация. Количество репликонов на одну интерфазную хромосому у эукариот может достигать 100 и более. В клетке млекопитающих может быть 20 – 30 тыс. репликонов, у человека – примерно 50 тыс. При фиксированной скорости роста цепи (у эукариот – 100 нуклеотидов в секунду) множественная инициация обеспечивает большую скорость процесса и снижение времени, необходимого для дупликации протяженных участков хромосом, т.е. у эукариот осуществляется полирепликонная репликация. (рис. 21) Репликон содержит все необходимые гены и регуляторные последовательности, которые обеспечивают репликацию. Каждый репликон в процессе клеточного деления активируется один раз. Репликация контролируется на стадии инициации. Если процесс удвоения начался он будет продолжаться до тех пор, пока весь репликон не будет удвоен. У прокариот вся ДНК является одним репликоном.
Рис.21. Репликация хромосомной ДНК эукариот. Репликация идет в двух направлениях из разных точек начала репликации (Ori) с образованием пузырьков. «Пузырь» или «глаз» это область реплицированной ДНК внутри нереплицированной. (А. С. Коничев, Г. А. Севастьянова, 2005, с. 213)
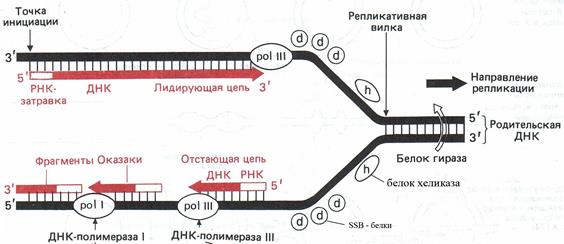
Ферменты, участвующие в процессе репликации, объединены в мультиферментативный комплекс. В репликации ДНК у прокариот участвует 15 ферментов, а у эукариот – более 30, т.е. репликация – это архисложный и суперточный многоступенчатый ферментативный процесс. В состав ферментативных комплексов входят следующие ферменты: 1) ДНК – полимеразы (I, III), катализируют комплементарное копирование, т.е. отвечают за рост дочерней цепи. (рис. 22) Прокариоты реплицируются со скоростью 1000 нуклеотидов в секунду, а эукариоты - 100 нуклеотидов в секунду. Пониженная скорость синтеза у эукариот связана с затрудненной диссоциацией гистоновых белков, которые необходимо удалить для продвижения ДНК-полимеразы в репликативной вилке вдоль цепи ДНК. 2) ДНК - праймаза. ДНК – полимеразы могут удлинять полинуклеотидную цепь присоединяясь к уже имеющимся нуклеотидам. Поэтому, чтобы ДНК – полимераза смогла начать синтез ДНК, ей необходима затравка или праймер (от. англ. primer – затравка). ДНК – праймаза синтезирует такую затравку, которая затем замещается сегментами ДНК. (рис. 22). 3) ДНК – лигаза, соединяет фрагменты Оказаки друг с другом за счет образования фосфодиэфирной связи. 4) ДНК – хеликаза, расплетает спираль ДНК, разрывает водородные связи между ними. В результате образуются две одиночные разнонаправленные ветви ДНК (рис.22). 5) SSB – белки, связываются с одноцепочечной ДНК и стабилизируют её, т.е. они создают условия для комплементарного спаривания. Репликация ДНК начинается не в любой случайной точке молекулы, а в специфических местах, называемых областью (точками) начала репликации (Ori). Они имеют определенные последовательности нуклеотидов, что облегчает разделение цепей (рис.21). В результате инициации репликации в точке Ori образуются одна или две репликативные вилки – места разделения материнских цепей ДНК. Процесс копирования продолжается до тех пор, пока ДНК полностью не удвоится или пока репликативные вилки двух соседних точек начала репликации не сольются. Точки начала репликации у эукариот разбросаны по хромосоме на расстоянии равном 20 000 пар нуклеотидов (рис.21).
Рис.22. Репликация ДНК (объяснение в тексте). (Б. Альбертс и др., 1994, т. 2, с. 82) Фермент – хеликаза – разрывает водородные связи, т.е. расплетает двойную цепь, образуя две разнонаправленные ветви ДНК (рис.22). Одноцепочечные участки связываются специальными SSВ-белками, которые выстраиваются снаружи каждой материнской цепи и оттягивают их друг от друга. Это делает азотистые основания доступными для связывания с комплементарными нуклеотидами. В месте схождения этих ветвей по направлению репликации ДНК располагается фермент ДНК-полимераза, который катализирует процесс и контролирует точность комплементарного синтеза. Особенностью работы данного фермента является его однонаправленность, т.е. построение дочерней цепи ДНК идет по направлению от 5' конца к 3'. На одной материнской цепи синтез дочерней ДНК идет непрерывно (лидирующая цепь). Она растет от 5' к 3' концу в направлении движения репликативной вилки и поэтому нуждается только в одном акте инициации. На другой материнской цепи синтез дочерней цепи идет в виде коротких фрагментов с обычной 5' - 3' полярностью и при помощи ферментов – лигаз происходит их сшивание в одну неперывную отстающую цепь. Поэтому для синтеза отстающей цепи требуется несколько актов (точек) инициации. Такой способ синтеза назван прерывистой репликацией. Фрагментные участки, синтезированные на отстающей цепи, в честь первооткрывателя названы фрагментами Оказаки. Они обнаружены у всех реплицирующихся ДНК, как у прокариот, так и у эукариот. Их длина соответствует 1000 – 2000 нуклеотидам у прокариот и 100 – 200 у эукариот. Таким образом, в результате репликации образуются 2 идентичные молекулы ДНК, в которых одна цепь материнская, другая вновь синтезированная. Такой способ репликации называют полуконсервативным. Предположение о таком способе репликации было сделано Дж. Уотсоном и Ф. Криком, а доказано в 1958г. М. Мезелсоном и Ф. Сталем. После репликации хроматин представляет собой систему из 2 декомпактизированных молекул ДНК, объединенных центромерой. В процессе репликации могут возникать ошибки, которые у прокариот и эукариот бывают с одной и той же частотой - одна на 108 -1010 нуклеотидов, т.е. в среднем 3 ошибки на геном. Это доказательство высокой точности и скоординированности процессов репликации. Ошибки репликации исправляются ДНК-полимеразой III («механизм корректорской правки») или системой репараций. 2. Репарация - это свойство ДНК восстанавливать свою целостность, т.е. исправлять повреждения. Передача наследственной информации в неискаженном виде важнейшее условие выживания как отдельного организма, так и вида в целом. Большинство изменений вредны для клетки, они либо приводят к мутациям, либо блокируют репликацию ДНК, либо вызывают гибель клетки. ДНК постоянно подвергается действию спонтанных (ошибки репликации, нарушение структуры нуклеотида и т.д.) и индуцированных (УФ – облучение, ионизирующая радиация, химические и биологические мутагены) факторов среды. В ходе эволюции выработалась система позволяющая исправлять нарушения в ДНК – система репарации ДНК. В результате её активности на 1000 повреждений ДНК только одно приводит к мутациям. Повреждение - любое изменение ДНК, которое вызывает отклонение от обычной двуцепочечной структуры: 1) появление одноцепочечных разрывов; 2) удаление одного из оснований, в результате чего его гомолог остается неспаренным; 3) замещение одного основания в комплементарной паре другим, неправильно спаренным с основанием-партнером; 4) появление ковалентных связей между основаниями одной цепи ДНК или между основаниями на противоположных цепях. Репарация может проходить до удвоения ДНК (дорепликативная репарация) и после удвоения ДНК (пострепликативная). В зависимости от характера мутагенов и степени повреждения ДНК в клетке идет световая (фотореактивация), темновая, SOS-репарация и др. Считают, что фотореактивация идет в клетке, если повреждения ДНК вызваны естественными условиями (физиологические особенности организма, обычные факторы среды, в том числе - ультрафиолетовые лучи). Восстановление целостности ДНК при этом, происходит с участием видимого света: репаративный фермент активируется квантами видимого света, соединяется с поврежденной ДНК, разъединяет пиримидиновые димеры нарушенного участка и восстанавливает целостность нити ДНК. Темновая репарация (эксцизионная) наблюдается после действия ионизирующей радиации, химических веществ и т.д. Она включает удаление поврежденного участка, восстановление нормальной структуры молекулы ДНК (рис.23). Для этого типа репарации необходима вторая комплементарная цепь ДНК. Темновая репарация многоступенчата, в ней участвует комплекс ферментов, а именно: 1)фермент, узнающий поврежденный участок цепи ДНК 2)ДНК – эндонуклеаза, делает разрыв в поврежденной цепи ДНК 3) экзонуклеаза удаляет измененную часть нити ДНК 4) ДНК – полимераза I синтезирует новый участок ДНК взамен удаленного 5)ДНК- лигаза сшивает конец старой нити ДНК с вновь синтезированной, т.е. замыкает два конца ДНК (рис.23). В темновой репарации у человека принимают участие 25 белков-ферментов. При больших повреждениях ДНК, которые угрожают жизни клеток, включается SOS-репарация. SOS-репарация была открыта в 1974 году. Такой тип репарации отмечают после действия больших доз ионизирующей радиации. Характерная черта SOS-репарации - неточность восстановления первичной структуры ДНК, в связи с чем она получила название репарации, склонной к ошибкам. Главная цель SOS-репарации сохранить жизнеспособность клетки. Нарушение в системе репарации могут приводить к преждевременному старению, развитию онкологических заболеваний, болезням аутоиммунной системы, гибели клетки или организма.
Функции ДНК Строение и свойства ДНК определяют ее основные функции: 1. Хранение генетической информации. ДНК находится в ядре и исключена из активных обменных процессов. 2. Передача генетической информации потомству происходит в процессе митоза и мейоза на основе репликации ДНК. 3. Запись генетической информации. Генетическая информация записана в виде ГЕНЕТИЧЕСКОГО или биохимического кода. 4. Контроль за обменом веществ в клетке
|
|||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-10; просмотров: 2128; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.227.13.119 (0.011 с.) |



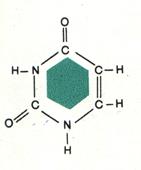







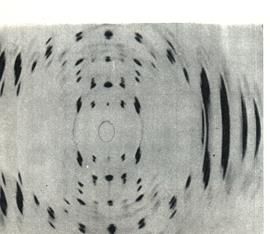


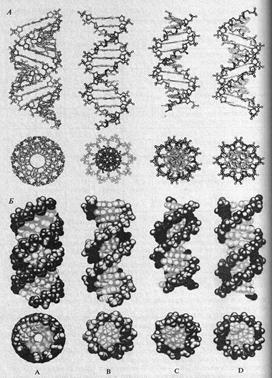
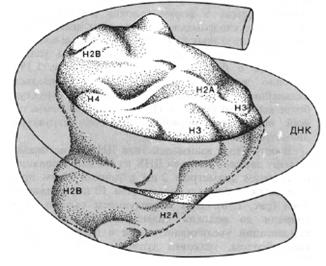
 Рис. 13. Модель нуклеосомного кора. Сегмент ДНК (146 пар оснований), обвивает белковый кор, делая вокруг него примерно 2 оборота (1¾). (С. Б. Бокуть и др., 2005, с. 52)
Рис. 13. Модель нуклеосомного кора. Сегмент ДНК (146 пар оснований), обвивает белковый кор, делая вокруг него примерно 2 оборота (1¾). (С. Б. Бокуть и др., 2005, с. 52)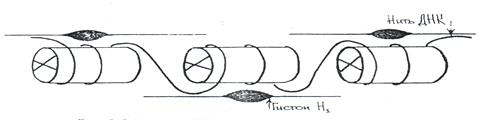







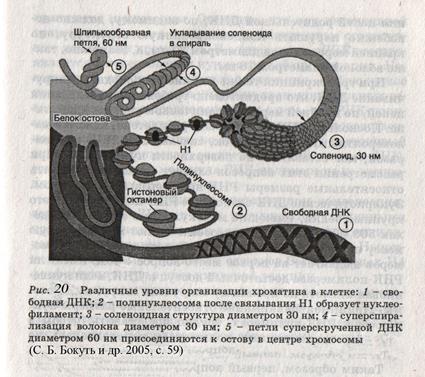 На этом этапе происходит объединение петель имеющих одинаковую организацию, образуются блоки или минидиски. В образовании одного минидиска участвуют примерно около 20 петель. Таким образом, за счет нескольких уровней компактизации длина ДНК сокращается примерно в 10000 раз. К онденсация хромосом из деконденсированного состояния - это не спирализация, а очень сложный комплекс компактизации, связанный не только с изменением их линейных размеров, но и с регуляцией их работы в процессе жизнедеятельности клетки. (Рис. 20)
На этом этапе происходит объединение петель имеющих одинаковую организацию, образуются блоки или минидиски. В образовании одного минидиска участвуют примерно около 20 петель. Таким образом, за счет нескольких уровней компактизации длина ДНК сокращается примерно в 10000 раз. К онденсация хромосом из деконденсированного состояния - это не спирализация, а очень сложный комплекс компактизации, связанный не только с изменением их линейных размеров, но и с регуляцией их работы в процессе жизнедеятельности клетки. (Рис. 20) 2. Репликация или самоудвоение ДНК: ДНК↔ДНК. Генетическая программа клеточных организмов записана в нуклеотидной последовательности ДНК. Для сохранения уникальных свойств организма необходимо точное воспроизведение этой последовательности в каждом последующем поколении. Во время деления клетки содержание ДНК должно удвоиться, чтобы каждая дочерняя клетка могла получить полный спектр ДНК, т.е. в любой делящейся соматической клетке человека должно быть скопировано 6,4*109 нуклеотидных пар. Процесс удвоения ДНК получил название репликации. Репликация относится к реакциям матричного синтеза. Во время репликации каждая из двух цепей ДНК служит матрицей для образования комплементарной (дочерней) цепи. Протекает она в S-период интерфазы клеточного цикла. Высокая надежность процесса репликации гарантирует практически безошибочную передачу генетической информации в ряду поколений. Пусковым сигналом для начала синтеза ДНК в S-периоде является так называемый S – фактор (специфические белки). Зная скорость репликации и длину хромосомы эукариот можно рассчитать время репликации, которое теоретически составляет несколько суток, а практически репликация осуществляется за 6 – 12 часов. Из этого следует, что репликация у эукариот одновременно начинается в нескольких местах на одной молекуле ДНК.
2. Репликация или самоудвоение ДНК: ДНК↔ДНК. Генетическая программа клеточных организмов записана в нуклеотидной последовательности ДНК. Для сохранения уникальных свойств организма необходимо точное воспроизведение этой последовательности в каждом последующем поколении. Во время деления клетки содержание ДНК должно удвоиться, чтобы каждая дочерняя клетка могла получить полный спектр ДНК, т.е. в любой делящейся соматической клетке человека должно быть скопировано 6,4*109 нуклеотидных пар. Процесс удвоения ДНК получил название репликации. Репликация относится к реакциям матричного синтеза. Во время репликации каждая из двух цепей ДНК служит матрицей для образования комплементарной (дочерней) цепи. Протекает она в S-период интерфазы клеточного цикла. Высокая надежность процесса репликации гарантирует практически безошибочную передачу генетической информации в ряду поколений. Пусковым сигналом для начала синтеза ДНК в S-периоде является так называемый S – фактор (специфические белки). Зная скорость репликации и длину хромосомы эукариот можно рассчитать время репликации, которое теоретически составляет несколько суток, а практически репликация осуществляется за 6 – 12 часов. Из этого следует, что репликация у эукариот одновременно начинается в нескольких местах на одной молекуле ДНК.