
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Строение гена про- и эукариотСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Гены прокариот состоят из двух основных элементов: регуляторной части и собственно кодирующей части(рис. 27). Регуляторная часть обеспечивает первые этапы реализации генетической информации, а кодирующая часть содержит информацию о структуре полипептида, тРНК, рРНК. У прокариот структурные гены, кодирующие белки одноо метаболического пути, часто бывают объединены и называются опероном. Так, например, в лактозном опероне E. coli содержится 3 структурных гена. Для биосинтеза аминокислоты гистидин требуется 9 ферментов и ее оперон содержит 9 структурных генов.
( А. С. Коничев, Г. А. Севастьянова, 2005, с. 157)
Гены эукариот имеют более сложное строение. В 1978г. У. Гильберт предположил: эукариотический геном состоит из модульных единиц, что позволяет «смешивать» и «сочетать» части. Онна основании анализа многих работ предложил модель мозаичного (интронно-экзонного) строения гена эукариот (28). Интроны – это некодирующие последовательности, они не входят в состав зрелых РНК. Экзоны – это последовательности участвующие в образовании зрелых РНК. Они могут быть как кодирующие так и некодирующие. Наследственная информация экзонов реализуется в синтезе определенных белков, а роль интронов до конца еще не выяснена. Возможное значение интронов: 1. Интроны снижают частоту мутаций, соотношение интронов и экзонов у человека 3:2. 2. Интроны поддерживают структуру ДНК, т.е. играют конститутивную роль. 3. Интроны необходимы для процесса созревания иРНК. Без интронов нарушен выход иРНК в цитоплазму. При введение в ядро искусственной иРНК без интронов, она остается в ядре и в цитоплазму не выходит. 4. В последние годы четко установлено, что некоторые интроны кодируют белки – ферменты, которые их вырезают. 5. Превращаются в малые ядерные РНК (мяРНК).
(А. С. Коничев, Г. А. Севастьянова, 2005 г., с. 157)
Гены высших организмов чаще оказываются прерывистыми, но есть и непрерывистые, например, гены интерферонов, гены гистонов. Степень прерывистости может быть различной – от одного интрона как у гена актина до нескольких десятков, как у гена коллагена(рис.29).
Рис. 29. Карты некоторых прерывистых генов. Жирные линии – экзоны, тонкие - интроны (А. С. Коничев, Г. А. Севастьянова, 2005 г., с. 158)
Длина интронов часто оказывается длинней экзонов: 5 – 20 тыс. и 1 тыс. соответственно. Прерывистость гена считалось достоянием эукариот. Но в 1983г. группа ВЕЗЕ (США) обнаружила их у некоторых археобактерий. Интроны содержаться во всех типах РНК, Интроны в составе иРНК вырезаются при участии мяРНП, которые образуют с интроном сплайсосому. При помощи сплайсосом, узнается начало и конец интрона, их концы соединяются в цепи РНК и интрон вырезается (рис.32). Эволюционное возникновение мозаичной (итрон – экзонной) структуры генов эукариот в настоящее время не находит объяснения. С точки зрения У. Гильберта появление интронов обеспечило возможность обмена экзонами между неродственными генами. В результате это привело к появлению белков с новыми функциями (гипотеза позднего возникновения интронов). По другой гипотезе интроны это эволюционные реликты, они были частью гигантских генов. Прокариоты являются эволюционным тупиком т.к. не содержат интронов.
Регуляция работы гена Гены функционируют в клетке не сами по себе, а входят в состав более сложной генной регуляторной системы. Количество структурных генов в разных оперонах различно. Участок ДНК, на на котором проходит считывание информации, называется единицей транскрипции (Рис. 27, 28). Он ограничен промотором (зона начала транскрипции) и терминатором (зона остановки транскрипции). 1. Промотор – это строго определенная нуклеотидная последовательность, которая узнается ферментом транскрипции – РНК – полимеразой. У E. coli промотор – это пара нуклеотидных последовательностей из 6-7 и 9 нуклеотидов каждая, отдельных друг от друга 25 нуклеотидами. Промотор выполняет следующие функции: а) это место присоединения РНК – полимеразы к молекуле ДНК б) последовательность оснований в промоторе определяет какая из цепей ДНК будет «смысловой», т.е. с какой цепи ДНК будет идти считывание информации (РНК – полимераза всегда двигается по цепи ДНК от 3' к 5' концу). У про- и эукариот последовательности промоторов разные. Это учитывается в генной инженерии, в случае встраивания в геном бактерии генов человека. Промоторы эукариот разнообразны по числу и строению элементов. Промотор эукариотического гена – это участок ДНК, на котором собираются белки транскрипции, узнающие свои сайты связывания и взаимодействующие друг с другом и с иРНК – полимеразой. В составе эукариотического гена имеются особые цис-действующие элементы регуляции - усилители или активаторы, глушители или угнетатели транскрипции. Они разнообразны по строению, положению и функциям. Они могут располагаться как на 5' - так и на 3 '- конце фрагмента ДНК, включающего ген, так и в составе интронов. 2. Оператор – это нетранскрибирумая последовательность нуклеотидов, участок связывания белка - репрессора. Он располагается в непосредственной близости к промотору или перекрывается с ним. У многих оперонов имеется не один, а несколько сайтов связывания с регуляторными белками, которые не обязательно располагаются рядом, а могут находиться по разные стороны от промотора. Поэтому сейчас принято говорить о сайтах связывания регуляторов. Связывание белка – репрессора с оператором либо создает стерические (пространственные) затруднения для связывания РНК-полимеразы с промотором, либо препятствует продвижению ее по смысловой цепи ДНК, и определяет точку начала транскрипции. Следует отметить, что ни промотор, ни оператор в РНК не транскрибируются и зоны промотора и оператора могут перекрываться. 3. Терминатор – участок молекулы ДНК, где заканчивается процесс транскрипции. Оператор и структурные гены образуют оперон. Именно так назвали эту структуру французские ученые Франсуа Жакоб и Жак Моно, которые первыми в 1959 -1961г. работая с бактериальными клетками предложили механизм регуляции работы гена или генной экспрессии. За эту работу в 1965г. они получили Нобелевскую премию. Как выяснили Жакоб и Моно работой оперона управляют гены – регуляторы. Они не входят в состав оперона, но являются необходимой частью регуляторной системы. Гены – регуляторы у прокариот находятся на той же хромосоме, что и оперон. У эукариот они могут располагаться далеко от промотора эукариотического гена и оказывать дистанционное влияние на его транскрипцию. Гены - регуляторы контролируют синтез белка – репрессора связывающегося с оператором. Синтез белков – репрессоров, как и всех белков, идет на рибосомах в цитоплазме. Транскрипция определяется белком-репрессором, который может закрывать оператор (репрессор активен) или открывать его (репрессор неактивен), т.е. возможны два варианта регуляции активности генов. I. Ген – регулятор отвечает за синтез активного белка – репрессора. Белок – репрессор имеет два активных центра: 1. центр связывания с оператором 2. центр связывания с субстратом. Под субстратом (индуктором) понимают любое вещество, информация о синтезе или распаде которого закодирована в данном опероне или гене. Это могут быть гормоны, аминокислоты, углеводы, питательные вещества, яды и т.д. Субстрата в клетке нет, поэтому активная форма белка – репрессора соединяется с оператором, т.е. оператор закрыт и через него не может пройти фермент РНК – полимераза, транскрипция не идет (рис.30). Открытие оператора идет с помощью субстрата (индуктора), поступающего в клетку. Индуктор взаимодействует с белком – репрессором, что приводит к изменению его конформации (пространственной структуры). У инактивированного белка репрессора резко снижается родство к зоне оператора и он отсоединяется от него. Оператор свободен и это позволяет РНК-полимеразе начать транскрипцию. Она продолжается до тех пор, пока в клетке есть субстрат, т.е. пока есть необходимость в продуктах данного оперона или гена (рис.31). При сокращении количества субстрата его уже не хватает на молекулы белка – репрессора и активный белок – репрессор присоединяется к оператору. Транскрипция прекращается. Следует отметить, что в клетке белок реперссор синтезируется постоянно и его количество строго определенное. Например, в клетке E. сoli находится около 10 молекул белка-репрессора, который регулирует работу лактозного оперона. II. Ген – регулятор отвечает за синтез неактивной формы белка – репрессора, т.е. он не может присоединится к оператору. Оператор свободен иРНК – полимераза свободно проходит к структурным генам. Оперон будет работать до тех пор, пока есть необходимость в продуктах данного оперона. Когда данный продукт клетке уже не нужен (он не расходуется в биохимических процессах и накапливается в клетке), субстрат взаимодействует с неактивным белком – репрессором, активирует его. Белок-репрессор закрывает оператор и выключает транскрипцию. Такой способ регуляции метаболизма в клетке чрезвычайно экономичен, т.к. клетка синтезирует продукт в таком количестве, которое необходимо для поддержания определенного уровня обменных процессов. При избытке конечного продукта данный метаболический путь выключается. То есть, мы видим взаимодействие между внутриклеточной средой и генетическим аппаратом для обеспечения тонкой регуляции клеточного метаболизма. У эукариот регуляция белкового синтеза еще сложнее и осуществляется на многих этапах от ДНК к белку. Но рассмотренные механизмы регуляции работы генов имеют место и у эукариот. Ж. Моно сказал: «Что хорошо и правильно для бактерий с генетической точки зрения, то правильно и для слона» Например, образование некоторых ферментов индуцируется присутствием их субстрата: 1. наличие в крови алкоголя индуцирует в клетках печени усиленный синтез фермента, разрушающего алкоголь – алкогольдегидрогеназы. 2. действие половых гормонов при формировании вторичных половых признаков также основано на усилении транскрипции определенных генов. 3. по такому типу работают гены железистых клеток, вырабатывающие секреты для жизнедеятельности организма. Если у бактерии на включение гена в работу требуется несколько минут, то у эукариот от нескольких часов до нескольких дней. Включение и работа генов и оперонов зависят от ряда факторов: 1. Специализации клетки 2. Физиологического состояния 3. Возраста клетки 4. Условий внешней среды 5. Пространственной структуры ДНК (изгибы, петли, сверхспирали и т.д.) 6. Степени метилирования генов. Показано, что гены материнских и отцовских хромомсом могут быть метилированы по – разному и это регулирует активность разных генов. Например, ген – индуцирующий образование опухолей. Если он передается потомству от отца, то транскрибируется только в сердце, а если от матери, то он вообще не экспрессируется. Исследования показали, что у самок этот ген метилирован, а у самцов – деметилирован. Любой из этих факторов может оказать существенное влияние на процесс считывания генетической информации.
|
||||
|
|
Последнее изменение этой страницы: 2016-12-10; просмотров: 8045; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.12.147.137 (0.012 с.) |

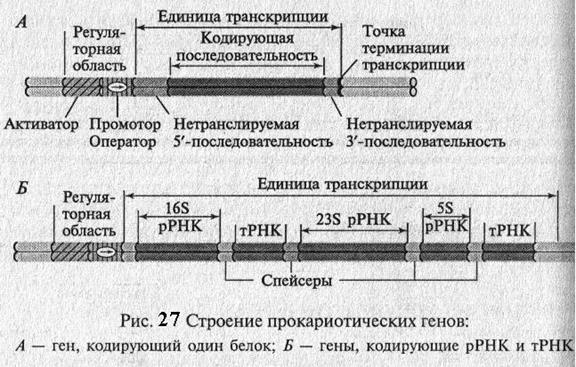 Гены, кодирующие белки, обычно содержат на 5'- и 3'- концах гена или оперона нетранслируемые последовательности (5' – НТП и 3' – НТП), которые играют важную роль в стабилизации иРНК. Гены тРНК и рРНК отделены друг от друга спейсерами (от англ. – spacer – распорка), т.е. последовательностями, которые вырезаются в ходе их созревания (процессинга)(рис. 27).
Гены, кодирующие белки, обычно содержат на 5'- и 3'- концах гена или оперона нетранслируемые последовательности (5' – НТП и 3' – НТП), которые играют важную роль в стабилизации иРНК. Гены тРНК и рРНК отделены друг от друга спейсерами (от англ. – spacer – распорка), т.е. последовательностями, которые вырезаются в ходе их созревания (процессинга)(рис. 27).




