Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Происхождение эукариотической клеткиСодержание книги Поиск на нашем сайте
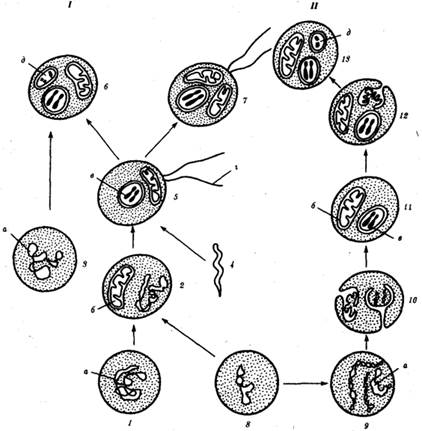
Ископаемые останки клеток эукариотического типа обнаружены в породах, возраст которых не превышает 1,0—1,4 млрд. лет. Более позднее возникновение, а также сходство в общих чертах их основных биохимических процессов (самоудвоение ДНК, синтез белка на рибосомах) заставляют думать о том, что эукариотические клетки произошли от предка, имевшего прокариотическое строение. Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой (рис. 1.4) основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней.
Рис. 1.4. Происхождение эукариотической клетки согласно симбиотической (I) и инвагинационной (II) гипотезам: 1 — анаэробный прокариот (клетка-хозяин), 2 — прокариоты, имеющие митохондрии, 3 — сине-зеленая водоросль (презумптивный хлоропласт), 4 — сиирохетообразная бактерия (презумпгивный жгутик), 5 — примитивный эукариот со жгутиком, 6 — растительная клетка, 7 — животная клетка со жгутиком, 8 — аэробный прокариот (презумптивная митохондрия), 9 — аэробный прокариот (клетка-родоначальница согласно гипотезе II), 10 — инвагинации клеточной оболочки, давшие ядро и митохондрии, 11 — примитивный эукариот 12 — впячивание клеточной оболочки, давшее хлоропласт, 13 — растительная клетка; а— ДНК прокариотической клетки, б — митохондрия, в — ядро эукариотической клетки, г — жгутик, д — хлоропласт Сходное происхождение предполагают для жгутиков, предками которых служили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза. Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили прокариотические синезеленые водоросли.
Серьезным доводом в пользу симбиотического происхождения митохондрий, центриолей и хлоропластов является то, что перечисленные органеллы имеют собственную ДНК. Вместе с тем белки бациллин и тубулин, из которых состоят жгутики и реснички соответственно современных прокариот и эукариот, имеют различное строение. У бактерий не найдено также структур со свойственной жгутикам, ресничкам, базальным тельцам и центриолям эукариотических клеток комбинацией микротрубочек: «9 + 2» или «9 + 0». Внутриклеточные мембраны гладкой и шероховатой цитоплазматической сети, пластинчатого комплекса, пузырьков и вакуолей рассматривают как производные наружной мембраны ядерной оболочки, которая способна образовывать впячивания. Центральным и трудным для ответа является вопрос о происхождении ядра. Предполагают, что оно также могло образоваться из симбионта-прокариота. Увеличение количества ядерной ДНК, во много раз превышающее в современной эукариотической клетке ее количество в митохондрий или хлоропласте, происходило, по-видимому, постепенно путем перемещения групп генов из геномов симбионтов. Нельзя исключить, однако, что ядерный геном формировался путем наращивания генома клетки-хозяина (без участия симбионтов). Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот (рис. 1.4). Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран. Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствует таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме эукариотической клетки.
История показала, что эволюционные возможности клеток эукариотического типа несравнимо выше, чем прокариотического. Ведущая роль здесь принадлежит ядерному геному эукариот, который во много раз превосходит по размерам геном прокариот. Количество генов у бактерии и в клетке человека, например, соотносится как 1: (100-1000). Важные отличия заключаются в диплоидности эукариотических клеток благодаря наличию в ядрах двух комплектов генов, а также в многократном повторении некоторых генов. Это расширяет масштабы мутационной изменчивости без угрозы резкого снижения жизнеспособности, эволюционно значимым следствием чего является образование резерва наследственной изменчивости. При переходе к эукариотическому типу усложняется механизм регуляции жизнедеятельности клетки, что на уровне генетического материала проявилось в увеличении относительного количества регуляторных генов, замене кольцевых «голых» молекул ДНК прокариот хромосомами, в которых ДНК соединена с белками. В итоге стало возможным считывать биологическую информацию по частям с разных групп генов в разном их сочетании в различных типах клеток и в разное время. В бактериальной клетке, напротив, одновременно считывается до 80—100% информации генома. В клетках взрослого человека в разных его органах транскрибируется от 8—10% (печень, почка) до 44% (головной мозг) информации. Использованию биологической информации частями принадлежит исключительная роль в эволюции многоклеточных организмов, так как именно это позволяет разным группам клеток специализироваться по различным функциональным направлениям. Большое значение при переходе к многоклеточности имело наличие у эукариотических клеток эластичной оболочки, что необходимо для образования устойчивых клеточных комплексов. Среди цитофизиологических особенностей эукариот, увеличивающих их эволюционные возможности, необходимо назвать аэробное дыхание, которое также послужило предпосылкой для развития многоклеточных форм. Интересно, что сами эукариотические клетки появились на Земле после того, как концентрация O2 в атмосфере достигла 1% (точка Пастера). Названная концентрация является необходимым условием аэробного дыхания. В условиях усложнения генетического аппарата эукариот, увеличения суммарного количества ДНК и распределения ее по хромосомам трудно переоценить значение возникновения в эволюции митоза как механизма воспроизведения в поколениях генетически сходных клеток. Появление вследствие эволюционных преобразований митоза такого способа деления клеток, как мейоз, дающего возможность сохранить постоянство хромосом в ряду поколений, наилучшим образом решило проблему размножения многоклеточных организмов. Связанный с мейозом переход к половому размножению усилил эволюционную роль комбинативной изменчивости, способствовал увеличению скорости эволюции. Благодаря отмеченным особенностям за 1 млрд. лет эволюции эукариотический тип клеточной организации дал широкое разнообразие живых форм от одноклеточных простейших до млекопитающих и человека.
8. Существование клетки зависит от выполнения ею ряда обязательных условий. К ним относятся отграничение от окружающей среды и вместе с тем обмен веществами с этой средой. На основе биохимических механизмов внутри клетки происходят реакции диссимиляции и ассимиляции, образуются химические соединения для выполнения тех или иных функций. В процессе жизнедеятельности возникают вещества, которые подлежат удалению. Приобретение клеткой способности к активному движению облегчает задачу поиска пищи и избегания опасных ситуаций. Сохранение жизни во времени зависит от способности клеток к делению. В ходе эволюции совершенствование жизненно важных функций происходит путем их дифференциации, т.е. обособления. Нередко такое обособление связано с возникновением специальных структур. У одноклеточных организмов, например у инфузории, это проявляется в приобретении некоторыми внутриклеточными структурами специализации (см. рис. 2.2). Так, пищеварительные вакуоли обеспечивают переваривание поступающих извне веществ с утилизацией клеткой необходимых химических соединений и выведением вовне непереваренных остатков. Функция сократительных вакуолей состоит в регулировании водного баланса, ресничек — в обеспечении двигательной активности.
Названная закономерность, проявляющаяся в разделении и специализации функций и структур, представляет собой одно из всеобщих свойств жизни. Возникновение среди живых форм многоклеточных организмов, с которыми связано прогрессивное направление эволюции, является логичным развитием этого свойства. В таких организмах усиление жизненной силы благодаря многократному повторению клеточных механизмов сочетается с широчайшим размахом разделения функций, их совершенствованием, образованием разнообразных специализированных структур — органов и их систем. Переход к многоклеточности — одновременно и новое качественное состояние жизни, для которого характерно ускорение эволюционных преобразований на основе более полного использования резерва наследственной изменчивости. Это обусловлено, во-первых, объединением у многоклеточных организмов полового процесса и размножения в единое целое — половое размножение (см. гл. 5). Во-вторых, хотя цикл индивидуального развития имеют все живые формы, включая вирусы, только у многоклеточных организмов выделяется эмбриональный период. Значение названного периода заключается в том, что, с одной стороны, в нем отражен весь длительный процесс исторического развития данного биологического вида, с другой — именно путем изменений в ходе эмбриогенеза происходят эволюционные изменения (см. § 13.2). Отмеченные особенности многоклеточной организации живых существ сделали их основой дальнейшей прогрессивной эволюции. Эволюционными предшественниками многоклеточных организмов были колониальные формы простейших организмов (см. § 13.1). Наиболее ранние ископаемые останки многоклеточных животных имеют возраст около 700 млн. лет. Палеонтологическая летопись свидетельствует о том, что многоклеточные организмы возникали в ходе эволюции от одноклеточных эукариот независимо не менее 17 раз. Из ныне существующих многоклеточных животных губки ведут свою родословную от одного предка, тогда как все другие формы — от какого-то другого. В процессе исторического развития на планете возникло не менее 35 типов многоклеточных организмов. Из них до сих пор существует 26, будучи представленными более чем 2 млн. видов.
9. всё в методичке
10. Прокариоты и эукариоты Основные различия между прокариотами и эукариотами:
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы, что, правда, несколько искажает первоначальное значение этого термина), из которых многие отделены от цитоплазмы одной или несколькими мембранами (в прокариотических клетках внутренние органоиды, окруженные мембраной, встречаются редко). Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. Тип питания осмотрофный. Генетический материал прокариот представлен одной молекулой ДНК, замкнутой в кольцо, имеется только один репликон. В клетках отсутствуют органоиды, имеющие мембранное строение. В геноме могут присутствовать мобильные генетические элементы, а у некоторых прокариот (например, вольбахия) их содержится необычно много. Изучение бактерий привело к открытию горизонтального переноса генов, который был описан в Японии в 1959 г. Это процесс широко распространен среди прокариот, а также у некоторых эукариот. Открытие горизонтального переноса генов у прокариот заставило по другому взглянуть на эволюцию жизни. Ранее эволюционная теория базировалась на том, что виды не могут обмениваться наследственной информацией. Прокариоты могут обмениваться генами между собой непосредственно (конъюгация, трансформация) а также с помощью вирусов - бактериофагов (трансдукция).
16. Классификация мутаций Геномные мутации. В результате мутаций в ядре зиготы изменяется видовое число хромосом. Кариотип особи изучается на метафазных пластинках.Геномные мутации могут касаться всех хромосом (полиплоидия) или отдельных хромосом (анеуплоидия). В последнем случае может добавляться отдельная хромосома (трисомия) или вместо пары будет представлена одна хромосома (моногамия). Эти мутации редко оказываются жизнеспособными, чаще они приводят летальному исходу еще в процессе эмбриогенеза (спонтанные аборты), либо к рождению ребенка с нарушениями умственного и физического развития (врожденными пороками развития). Таковы синдромы анеуплоидии в виде моно- и трисомий по аутосомным и половым хромосомам. В частности, известный синдром Дауна обусловлен трисомией по 21-й паре хромосом. Синдром Дауна связан с нарушением ряда признаков - искаженные физические способности, умственная отсталость, выраженная от легкой дебильности до тяжелых форм идиотии. Частота данного заболевания в поколении 1 на 500-700 новорожденных. Структурные мутации. В этом случае в результате мутации изменяется структура хромосомы. Основными видами структурных мутаций хромосом являются разрывные и обменные аберрации. К разрывным аберрациям относятся разного рода фрагменты (разделение хромосомы на части), к обменным аберрациям относятся случаи, когда имеются два разрыва и хромосома на их основе преобразуется, - транслокации (перенос участка хромосомы в другую хромосому или внутри хромосомы), инверсии (поворот участка внутри хромосомы на 180°), внутренние делеции (потеря внутренних участков хромосом), кольца (замыкание в кольцо отделяющегося внутреннего участка). Структурные изменения могут образовываться перемещающимися элементами в виде участков ДНК, мобильных по своему положению в организации генома. Как правило, структурные мутации хромосом приводят к множественным дефектам развития. Так, при делеции короткого плеча 5-й хромосомы (нехватке концевого фрагмента) наблюдается заболевание, названное синдромом "кошачьего крика". Помимо болезненного крика ребенка, напоминающего кошачье мяуканье, обусловленного аномалиями развития гортани, для больных детей характерны тяжелая умственная отсталость, задержка роста и другие симптомы. Ряд транслокаций и инверсий передаются потомкам. Генные мутации. При повреждении или нарушениях в порядке или замене нуклеотидов, появлении внутренней дупликации или делеции в Молекуле ДНК возникают генные (точковые) мутации. Эти изменения отдельных генов часто приводят к тяжелым дегенеративным заболеваниям, в частности, многочисленным болезням обмена веществ через нарушения синтеза белков, ферментов. Примером может служить мутация, приводящая к появлению серповидноклеточной анемии — наследственного заболевания, как правило, приводящего детей и подростков к смерти. В этом случае в эритроцитах вместо нормального гемоглобина A содержится аномальный гемоглобин S. Аномалию вызывает мутация Другой пример генной мутации - наследственное заболевание фенилкетонурия. Заболевание характеризуется резко выраженной умственной отсталостью, развивающейся вследствие нарушения нормальных биохимических процессов в мозге из-за накопления в организме фенилаланина. В отличие от нормальных детей у детей, больных фенилкетонурией, фенилаланин образуется при распаде тканевых белков и при переваривании белковой пищи. Мутация в гене, кодирующем фермент фенилала-нин-4-гидроксилазу, приводит к блокированию реакции превращения фенилаланина в тирозин. Что касается вредных доминантных мутаций, то каждый из ее носителей специфически страдает от нарушений жизнеспособности. Таким образом, отрицательные доминантные гены прямо входят в состав генетического груза данного поколения из-за мутаций, которые возникают в зародышевых клетках родителей.
Механизмы возникновения: Мутации по типу замены азотистых оснований. Эти мутации происходят в силу ряда конкретных причин. Одной из них может быть возникающее случайно или под влиянием конкретных химических агентов изменение структуры основания, уже включенного в спираль ДНК. Если такая измененная форма основания остается не замеченной ферментами репарации, то при ближайшем цикле репликации она может присоединять к себе другой нуклеотид. Примером может служить дезаминирование цитозина, превращающегося в урацил самопроизвольно или под влиянием азотистой кислоты.Образующийся при этом урацил, не замеченный ферментом ДНК-гликозилазой, при репликации соединяется с аденином, который впоследствии присоединяет тимидиловый нуклеотид. В результате пара Ц—Г замещается в ДНК парой Т—А. Дезаминирование метилированного цито зина превращает его в тимин Тимидиловый нуклеотид, являясь естественным компонентом ДНК, не обнаруживается ферментами репарации как изменение и при следующей репликации присоединяет адениловый нуклеотид. В результате вместо пары Ц—Г в молекуле ДНК также появляется пара Т—А. Мутации со сдвигом рамки считывания. Этот тип мутаций составляет значительную долю спонтанных мутаций. Они происходят вследствие выпадения или вставки в нуклеотидную последовательность ДНК одной или нескольких пар комплементарных нуклеотидов. Большая часть изученных мутаций, вызывающих сдвиг рамки, обнаружена в последовательностях, состоящих из одинаковых нуклеотидов. Изменению числа нуклеотидных пар в цепи ДНК способствуют воздействия на генетический материал некоторых химических веществ, например акридиновых соединений. Мутации по типу инверсии нуклеотидных последовательностей в гене. Данный тип мутаций происходит вследствие поворота участка ДНК на 180°. Обычно этому предшествует образование молекулой ДНК петли, в пределах которой репликация идет в направлении, обратном правильному. В пределах инвертированного участка нарушается считывание информации, в результате изменяется аминокислотная последовательность белка. Эволюционное значение: Мутации являются материалом для естественного отбора. Отбор не приводит к возникновению новых признаков, благодаря естественному отбору виды не возникают, а исчезают. Единственным источником видообразования есть мутационная изменчивость. Мутации происходят во всех возможных направлениях. Способность к мутациям наступает периодически. 17. Репарация — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК в клетке или в результате воздействия физическими или химическими агентами. Типы: Прямая репарация Прямая репарация наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты, способные быстро (как правило, в одну стадию) устранять соответствующее повреждение, восстанавливая исходную структуру нуклеотидов. Так действует, например, O6-метилгуанин-ДНК-метилтрансфераза, которая снимает метильную группу с азотистого основания на один из собственных остатков цистеина. Эксцизионная репарация Эксцизионная репарация (англ. excision — вырезание) включает удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы. Пострепликативная репарация Tип репарации, имеющей место в тех случаях, когда процесс эксцизионной репарации недостаточен для полного исправления повреждения: после репликации с образованием ДНК, содержащей поврежденные участки, образуются одноцепочечные бреши, заполняемые в процессе гомологичной рекомбинации при помощи белка RecA. Пострепликативная репарация была открыта в клетках E.Coli, не способных выщеплять тиминовые димеры. Это единственный тип репарации, не имеющий этапа узнавания повреждения. 18.Классификация нуклеотидных последовательностей в геноме эукариот: 1.Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки. 2. Низкочастотные повторы – последовательности, повторяющиеся десятки раз. 3. Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов. 4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие (~ 10 пн) некодирующие последовательности, которые входят в состав прицентромерного гетерохроматина. 19. Свойства гена 1. дискретность — несмешиваемость генов; 2. стабильность — способность сохранять структуру; 3. лабильность — способность многократно мутировать; 4. множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм; 5. аллельность — в генотипе диплоидных организмов только две формы гена; 6. специфичность — каждый ген кодирует свой признак; 7. плейотропия — множественный эффект гена; 8. экспрессивность — степень выраженности гена в признаке; 9. пенетрантность — частота проявления гена в фенотипе; 10. амплификация — увеличение количества копий гена. Для эукариотической клетки характерно:
1. Наличие интронов и экзонов в молекуле ДНК.
2. Созревание и-РНК - вырезание интронов и сшивка экзонов.
3. Наличие регуляторных элементов, регулирующих транскрипцию, таких как: а) промоторы - 3 вида, на каждый из которых садится специфическая полимераза. Pol I реплицирует рибосомные гены, Pol II - структурные гены белков, Pol III - гены, кодирующие небольшие РНК. Промотор Pol I и Pol II находятся перед участком инициации транскрипции, промотор Pol III - в рамках структурного гена; б) модуляторы - последовательности ДНК, усиливающие уровень транскрипции; в) усилители - последовательности, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена и состояния начальной точки синтеза РНК; г) терминаторы - специфические последовательности, прекращающие и трансляцию, и транскрипцию.
Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. Генетический материал прокариот представлен одной молекулой ДНК, замкнутой в кольцо, имеется только один репликон. В геноме могут присутствовать мобильные генетические элементы, а у некоторых прокариот (например, вольбахия) их содержится необычно много. Изучение бактерий привело к открытию горизонтального переноса генов. Открытие горизонтального переноса генов у прокариот заставило по другому взглянуть на эволюцию жизни. Ранее эволюционная теория базировалась на том, что виды не могут обмениваться наследственной информацией. Прокариоты могут обмениваться генами между собой непосредственно (конъюгация, трансформация) а также с помощью вирусов - бактериофагов (трансдукция).
20. Этапы реализации генетической информации в клетке Транскрипция является первым этапом общего переноса генетической информации и представляет собой процесс биосинтеза молекул РНК по программе ДНК. В качестве продуктов транскрипции (транскриптов) можно рассматривать все типы молекул РНК, участвующих в биосинтезе белков в клетке, — матричные (информационные) РНК (мРНК, или иРНК), рибосомные РНК (рРНК), транспортные РНК (тРНК), малые ядерные РНК (мРНК). Транскрипция подразделяется на три основные стадии — инициацию (начало синтеза РНК), элонгацию (удлинение полинуклеотидной цепочки) и терминацию (окончание процесса). Трансляция как очередной этап реализации генетической информации заключается в синтезе полипептида на рибосоме, при котором в качестве матрицы используется молекула мРНК (считывание информации в направлении 5' → 3'). Следует заметить, что в клетках прокариот, не имеющих настоящего ядра с оболочкой, хромосомный генетический материал (ДНК) практически находится в цитоплазме, что определяет непрерывный характер взаимосвязи процессов транскрипции и трансляции. Посттрансляционная модификация полипептида представляет собой завершающий этап реализации генетической информации в клетке, приводящий к превращению синтезированного полипептида в функционально активную молекулу белка. При этом первичный полипептид может претерпевать процессинг, состоящий в ферментативном удалении инициирующих аминокислот, отщеплении других (ненужных) аминокислотных остатков и в химической модификации отдельных аминокислот. Затем происходит процесс сворачивания линейной структуры полипептида за счет образования дополнительных связей между отдельными аминокислотами и формирование вторичной структуры белковой молекулы. На этой основе формируется еще более сложная третичная структура молекулы.
41. Макроэволюция. Её отношение с микроэволюцией.
Макроэволюцинадвидовая эволюция, в результате которой формируются более крупные таксоны (семейства, отряды, классы, типы). Она не имеет спецефических механизмов и осуществляется аналагично механизмам микроэволюции. А.Н. Северцев и И.И. Шмальгаузен установили 2 главных направления эволюционного процесса: биологическиц прогресс, биологический регресс. Биологический прогресс – характеризуется расширением ареала; увеличением численности вида; образованием новых популяций и систематических едениц; преобладание рождаемости над смертностью. Например: ареал зайца-русака расширился и образовалось 20 новых подвидов. Биологический регресс – сужение ареала; уменьшение численности вида; сокращение числа популяций и уменьшение систематических едениц; преобладание смертности над рождаемостью. Это приводит к сокращению числа видов в роде, числа родов в семействе, семейства в отряде и т.д. Часть видов, родов, семейств вымирают полностью. Например: снижение численности хвощей и плаунов. На грани вымирания находится уссурийский тигр.
|
||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-10; просмотров: 562; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.214.244 (0.019 с.) |