Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Органічні сполуки -вуглецевмісні речовини живих організмівСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Клітини побудовані із специфічного і водночас обмеженого набору однакових для усіх видів живих істот великих молекул, що містять вуглець. Завдяки малим розмірам і вмісту на зовнішній оболонці чотирьох електронів атом вуглецю може утворювати чотири міцні ковалентні зв'язки з іншими атомами. Важлива здатність атомів вуглецю з'єднуватись один з одним у ланцюги, кільця й утворювати великі і складні молекули. Основні групи цих молекул є відносно простими вуглеводами, жирними кислотами, амінокислотами і нуклеотидами. Вуглеводи є найважливішим джерелом енергії для клітин, вони запасають її, створюють резервні полісахариди. Головна функція жирних кислот - утворення клітинних мембран і участь в енергетичному обміні. Полімери, побудовані з амінокислот, представлені різноманітними і багатофункціональними молекулами білків. Нуклеотиди відіграють головну роль в акумуляції і перенесенні енергії (АТФ, НАД), однак основне значення полягає в тому, що вони є субодиницями інформаційних молекул РНК і ДНК. Білки. Клітини значною мірою складаються з білків, на яких припадає більше половини маси сухої речовини клітини. Білки визначають структуру і форму клітини; крім того, вони є рецепторами молекулярного розпізнавання і каталізу. Білки безпосередньо беруть участь у процесах обміну речовин і виконанні функцій клітиною. Білки побудовані з 20 різних амінокислот, кожна з яких має хімічну індивідуальність. Комбінації з 20 амінокислот можуть утворювати незліченну кількість різних за структурою і функціями білків. Білкові молекули утворюються за допомогою пептидных зв 'язків між амінокислотами. Кілька амінокислот, об'єднаних пептидними зв'язками, називають поліпептидами. Прості білки утворені тільки амінокислотами. До складу складних білків можуть входити ліпіди (ліпопроте'їди), вуглеводи (глікопротеїди). Багато ферментів містять в активному центрі речовини небілкової природи (коферменти). Рівні структурної організації білків наведені на. Амінокислотна послідовність білкової молекули визначає її просторову структуру. Структура полі-пептидного ланцюга стабілізується нековалентними взаємодіями між її частинами. Амінокислоти групуються всередині молекули, а виникнення локальних водневих зв'язків між пептидними групами призводить до утворення a-спіралей.
Рівні структурної організації білків: а - первинна структура; б - вторинна структура; в - третинна структура; г - четвертинна структура. Класифікація білків за їх функціями
У залежності від форми, білки можуть бути фібрилярними і глобулярними. Фібрилярні - довгі, видовжені, досить стабільні, погано розчинні у воді. Деякі мають здатність до скорочення, наприклад, актин, міозин. Більшості білків властива глобулярна структура. Вони мають вигляд сфери і добре розчинні у воді, наприклад, гемоглобін, альбумін, більшість ферментів. Функції білків. Функції білка визначаються хімічною будовою і фізико-хімічними властивостями поверхні. Специфічні місця на поверхні білка або всередині, утворені закономірно розташованими амінокислотними залишками, формують центри специфічного зв'язування інших речовин і визначають функцію того чи іншого білка. Основні функції білків: а) каталітична - ферменти в тисячі разів прискорюють хімічну модифікацію молекул-субстратів, забезпечують усі основні функції клітини; б) регуляторна - гормони білкової природи беруть участь у регуляції і координації багатьох метаболічних і фізіологічних процесів живих організмів; в) структурна - білки забезпечують утворення всіх елементів клітин і організму: органел, мембран, тканин, органів, а також структурний зв'язок між ними; г) захисна - білки захищають організм, створюють його захисні покриви, оболонки органів і клітин, утворюють антитіла, регулюють рН; д) рецепторна -білки розпізнають сигнали, що надходять із зовнішнього середовища, перетворюють їх і передають у необхідний відділ організму; є) транспортна функція полягає у здатності окремих білків переносити речовини до місця використання. Наприклад, переносник різних речовин через біомембрани, а також цитоскелет - гемоглобін; ж) рухова функція забезпечується м'язовими білками: актином, міозином та іншими, з яких складаються м'язові тканини; з) енергетична роль зв'язана з можливістю використання органічних молекул білків в енергетичному обміні. В результаті їх зруйнування утворюється АТФ. Нуклеїнові кислоти. Нуклеїнові кислоти - унікальні молекули, необхідні кожній клітині для збереження і передачі генетичної інформації. Нуклеїнові кислоти забезпечують процеси синтезу білків, а цим, у свою чергу, визначається характер обміну речовин, закономірності росту й розвитку, явища спадковості й мінливості. Є дві основні групи нуклеїнових кислот: ДНК і РНК. Вони відрізняються хімічною будовою і біологічними властивостями. Вуглеводи. Вуглеводи - це органічні речовини, що мають загальну формулу Сх(Н20)у, де х і у можуть мати різні значення. їх поділяють на три основні класи: моносахариди, дисахариди, полісахариди. Глюкоза є найбільш важливим моносахаридом у живих системах, ключовою ланкою енергетичного обміну і структурним мономером полісахаридів. Інші важливі моносахариди: фруктоза, галактоза, рибо-за та ін. Моносахариди добре розчинні у воді й дифундують у цитоплазму через клітинні мембрани. Основна функція - джерело енергії для різних метаболічних процесів. Внаслідок послідовних реакцій окиснюван-ня гексози перетворюються в остаточному підсумку на СО і НО. Сумарне рівняння реакції: С6Н12О6 + 6О2 → 6СО2 + 6Н2О + енергія Енергія, що вивільняється, генерується і запасається у вигляді двох сполук - АТФ і НАДФ. Моносахариди можуть шляхом трансамінування перетворюватися в деякі амінокислоти. Вони також утворюють субстрати для синтезу жирів. Найбільш важливими дисахаридами є мальтоза (утворена двома молекулами глюкози), сахароза (складається з глюкози і фруктози) і лактоза (складається з глюкози і галактози). їх основна функція - джерело енергії для клітини. Полісахариди побудовані з повторюваних молекул глюкози. У клітинах тварин це глікоген, у рослинних - крохмаль. Основна функція - запасають велику кількість енергії. Деякі полісахариди зв'язуються з білками й утворюють глікопротеїни. Ліпіди. Це нерозчинні у воді органічні речовини клітини, які можна вилучити тільки органічними розчинниками. Хімічно дуже різноманітна група, але практично всі вони є складними ефірами жирних кислот і спирту. Велика частина ліпідів клітини - це складні ефіри органічних кислот і спирту гліцеролу (гліцерину). Усі ліпіди є висококалорійним джерелом енергії клітини. Основним компонентом багатьох видів ліпідів є жирні кислоти. Це карбонові кислоти з довгими вуглеводневими "хвостами", наприклад, пальмітинова кислота: сн3-сн2-сн2-сн2-сн2сн2-сн2-сн2-сн2-сн2-сн2-сн2-сн2сн2-сн2-соон Різні види жирних кислот відрізняються кількістю вуглецевих ланок і ненасичених зв'язків. Жирні кислоти, зв'язуючись із гліцерином, утворюють три-ацилгліцероли (тригліцериди) й у такому вигляді зберігаються в якості енергетичного резерву (жир). У фосфоліпідах дві ОН-групи зв'язані з жирними кислотами, а одна ОН-група - з фосфатом. Жирні кислоти і фосфоліпіди мають гідрофобний хвостик і полярну голівку. Найважливіша функція жирних кислот - участь у побудові клітинних мембран. У воді вони утворюють ліпідні бішари, що са-моорганізовуються і складають основу всіх мембран клітини. 2. Морфологія клітини. Структурні компоненти цитоплазми і ядра. Цитоплазма і цитоскелет. Циклоз Цитоплазма складає основну масу клітини — це весь її внутрішній вміст, за винятком ядра. Містить 75-85 % води, 15-25 % білків і багато інших речовин, але в менших кількостях. При вивченні клітини за допомогою світлового мікроскопа цитоплазма є гомогенною, безбарвною, прозорою, в'язкою рідиною. Проте електронний мікроскоп дозволив виявити складну багатокомпонентну, поліфункціональ-ну, високовпорядковану структуру цитоплазми. Цитоплазма складається із цшпозолю (цитоплазматичний матрикс), внутрішньоклітинних органел і включень. Цитозоль. Цитозоль становить більшу частину цитоплазми (55 % від загального об'єму клітин), що не містить органел. Це колоїд, який складається зі складної суміші розчинених у воді органічних макромолекул - білків, жирів, вуглеводів та неорганічних речовин. Містить до 10000 різних видів білків, головним чином ферментів.
Клітина еукаріотів: 1 - цитозоль. Хімічний склад і властивості цитозолю. У цито-золі знаходяться неорганічні (вода, солі, гази) і органічні речовини. Неорганічні речовини. Вода є основною складовою частиною цитозолю. У середньому в клітинах міститься близько 75 % води. Завдяки своїм властивостям водне середовище забезпечує майже всі життєві процеси в клітинах. Зокрема, вода має такі властивості: 1) розчинник для речовин у клітині, внаслідок чого багато з них іонізуються водою, що полегшує хімічні реакції; 2) сприяє пересуванню речовин у клітині, із клітини в клітину в розчиненому стані; 3) ефективний термостабілізатор і зберігає тепло, утворене клітиною; 4) забезпечує постійний броунівський рух молекул. Солі становлять 1-2 % цитозолю. У водному середовищі вони утворюють іони. Більшість солей клітин - це карбонати, бікарбонати, фосфати, сульфати і хлориди солей натрію, калію, кальцію, магнію та заліза. У першу чергу, вони відіграють істотну роль у підтримці осмотичності і кислотності цитозолю. Багато з них беруть участь у біологічних процесах і входять до складу деяких білків. Гази. У клітинах наявні кисень, вуглекислий газ, азот і аміак. Кисень і азот надходять з атмосфери шляхом дифузії. Вуглекислий газ і аміак утворюються в клітині в результаті обміну речовин. CO утворюється як кінцевий продукт при окисних реакціях і постійно видаляється з клітин. Азот - інертний газ, він не бере участі в клітинних реакціях. Органічні речовини становлять 20-25 % від маси живої клітини. Основними групами цих речовин є: білки, жири, вуглеводи і нуклеїнові кислоти. Вони, насамперед, забезпечують специфіку будови і функції клітин, є енергетичними субстратами окис-нювання, утворюють запасні речовини тощо. Фізичні властивості цитозолю. Колоїдний вміст може переходити з рідкого стану - золю - у більш твердий - гель. Зміни в колоїдному стані пов'язані з різним розподілом колоїдних частинок у цитозолі. Перехід з одного стану в інший називають фазовим переходом. У стані золю його частинки розподілені менш випадково й рівномірно, що забезпечує чіткий рух молекул. У стані гелю частки утворюють агрегати між собою і з водою, що призводить до зв'язування вільної води і втрати руху цитоплазми. Рухливість молекул значно зменшується. Це означає, що в місцях "твердого" цитозолю швидкість обміну речовин обмежена, але в ділянках "рідкого" цитозолю спостерігаються максимальні швидкості біохімічних процесів. Перехід ділянок цитоплазми зі стану гелю у золь і навпаки зумовлює циклоз - рух ділянок цитоплазми. Цей процес, наприклад, лежить в основі формування псевдоподій у амеб і лейкоцитів. У цитозолі відбувається постійний броунівський рух молекул, постійне їх зіткнення, що зумовлює високу швидкість метаболічних реакцій. Колоїдний стан цитозолю забезпечує об'єм і форму клітин, а за допомогою хімічних буферів підтримується сталість рН. Броунівський рух молекул залежить від стану цитозолю: чим він більш "рідкий" - тим інтенсивні-ший рух молекул. Підвищення температури також призводить до збільшення інтенсивності руху і прискорення біохімічних реакцій. Броунівський рух забезпечується тепловим рухом молекул. При цьому кожна молекула робить обертово-поступальні рухи. Це забезпечує часте зіткнення молекул, наприклад, кожна молекула цитозолю має приблизно 1 млн. зіткнень за секунду. Таким чином, броунівський рух лежить в основі й є необхідною умовою перебігу всіх біохімічних реакцій обміну речовин. Біологічні властивості цитозолю. Хімічний склад і фізичний стан зумовлюють біологічні властивості цитозолю, що служать структурній цілісності і функціональній активності клітин. Насамперед -це підтримка метаболізму. Цитозоль - це середовище, де перебігають одночасно тисячі біохімічних реакцій. Вважається, що близько 70 % реакцій клітинного метаболізму відбувається в цитозолі, що містить тисячі різновидів ферментів. Це реакції гліколізу, глюконеогенезу, синтезу білків, жирних кислот, амінокислот, нуклеотидів та інші. На рибосомах у цитозолі синтезується багато білків, які використовуються клітиною для власних потреб. Рибосоми, зв'язані з ЕПС, утворюють білки на "експорт". Функції органел клітини забезпечуються постійним, необхідним для них оточенням цитозолю. З цитозолю органели одержують необхідні речовини і викидають у нього відходи. Цитозоль бере участь у процесі підтримки гомеостазу клітини. Реакції, що відбуваються у цитозолі, забезпечують сталість складу клітини та її структурної організації. У цитозолі постійно підтримується концентрація води, газів, субстратів хімічних реакцій, рН. Ці умови необхідні для перебігу біохімічних та фізіологічних процесів. Внаслідок постійного синтезу молекул (білків, амінокислот, нуклеотидів, вуглеводів, жирів та ін.) можливий обмін ушкоджених молекул на нові, синтезовані. Це стосується і постійної підтримки структури і складу всіх органел. У цитозолі присутні нелізосомальні протеази, які перетравлюють дефектні білки з низькою тривалістю життя. Цитозоль є резервуаром різних субстратів (амінокислот, нуклеотидів, глюкози й інших), які постійно використовуються в обміні речовин для утворення нових структур або їх відновлення. Цитозоль забезпечує ріст і диференціювання клітини. Після поділу клітини мають малий розмір і слабко диференційовані. Ріст їх насамперед пов'язаний із синтезом і накопиченням необхідних органічних речовин, більшість яких утворюються в цитозолі. Ці речовини збільшують об'єм клітини, а також використовуються для формування або росту органел. У процесі розвитку клітин з'являються специфічні органели, змінюється форма клітин, вони поступово набувають рис та властивостей клітин-попередників. Таким чином, цитозоль є одним з основних компонентів клітини.
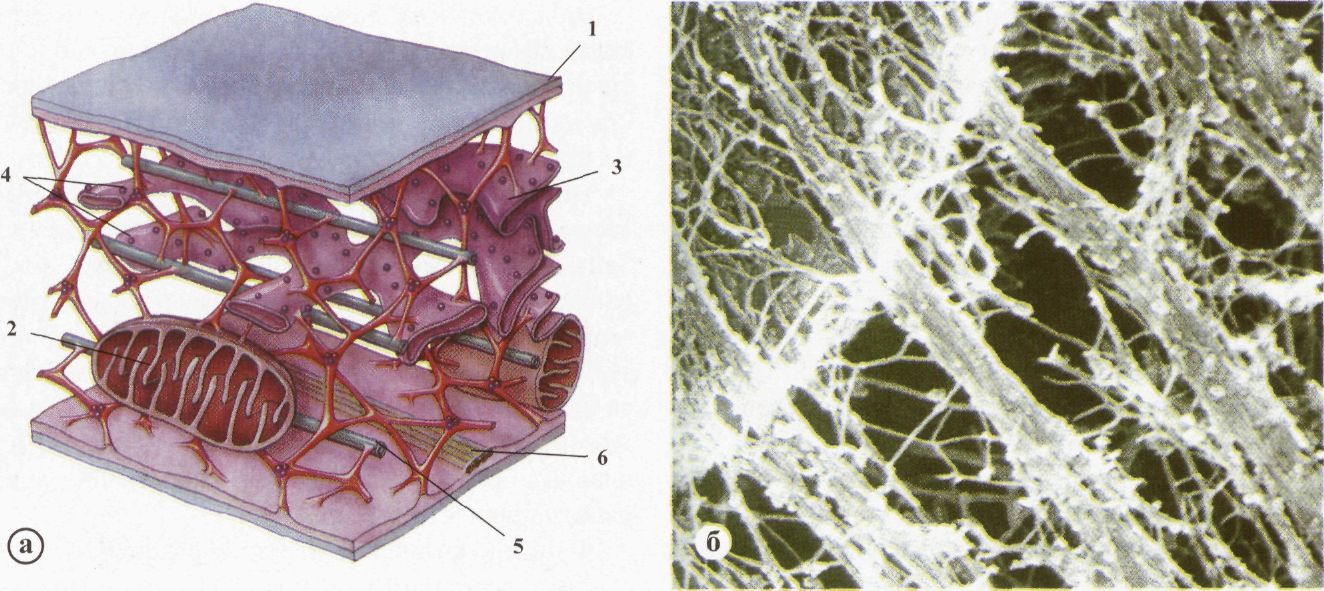
Цитоскелет (а - схематичне зображення; б - мікрофотографія): 1 - клітинна мембрана; 2 - мітохондрія; 3 - комплекс Гольджі; 4 - рибосоми; 5 - мікротрубочки; 6 - мікрофіламенти. Цитоскелет. Цитоскелет - це сітка білкових фібрил і мікротрубочок, що вкривають зсередини цитоплазматичну мембрану і пронизують внутрішній простір клітини (рис. 1. 28). Він характерний для всіх еукаріотичних клітин, а також є основним компонентом ворсинок і джгутиків найпростіших, хвостика сперматозоїдів, веретена поділу клітин. Цитоскелет складається з трьох типів структур: 1) мікротрубочки (найтовстіші), утворені кількома білковими фібрилами, які містять глобулярний білок - тубулін; 2) мікрофіламенти (найтонші), що мають здатність скорочуватися, утворюються глобулярним білком — актином; 3) проміжні філаменти (комбінація кількох мікрофіламентів). Фібрили цитоскелета можуть за необхідності згруповуватися з мономерів білків і розпадатися після виконання функції. Мають здатність до скорочення і руху. У клітині фібрили взаємодіють між собою за участі допоміжних білків. Вони вкривають з внутрішнього боку цитоплазматичну мембрану і пронизують внутрішній простір клітини. Цим досягається стабільність форми й об'єму клітини, а також можливість зміни форми, руху органел і клітини. Функції цитоскелета. 1. Підтримка об'єму і форми клітин. Основну роль у цьому відіграє фібрилярна сітка, що вкриває зсередини мембрану (кортекс). Ця сітка спеціальним білком (анкерін) прикріплена до цитолеми. До цієї сітки приєднані нитки мікрофіламєнтів і мікротрубочок, що значною мірою стабілізує форму клітини. 2. Зміна форми клітин. Система білкових фібрил здатна до скорочення або розтягування. За рахунок цього може відбуватися зміна форми клітин (наприклад, формування псевдоподій у лейкоцитах). 3. Пересування органел і транспортних везикул. Фібрили цитоскелета прикріплені до клітинних органел. Це стабілізує їхнє положення в цитоплазмі. З іншого боку, зміна довжини фібрил призводить до переміщення клітинних структур. 4. Утворення мультиферментних компонентів. У місцях переплетення кількох фібрил цитоскелета створюються сприятливі умови для розміщення комплексу ферментативних білків. Це забезпечує структурнуєдність ферментів та певний метаболічний процес. 5. Завдяки наявності щільної сітки мікрофібрил цитозоль набуває певної структури, що сприяє координованому розміщенню комплексів ферментів. Цим досягається інтеграція всієї цитоплазми - об'єднання в єдине ціле. 6. Утворення веретена поділу під час мітозу. Веретено поділу утворене сіткою мікротрубочок, що "збираються" за участі центріоль і чітко впорядковано розташовуються в цитозолі. 7. Утворення ворсинок і джгутиків у найпростіших. 8. Утворення міжклітинних контактів (десмосом). Десмосоми - структури цитоплазматичних мембран, що належать одночасно двом сусіднім клітинам. Зв'язування клітин відбувається завдяки мікрофіламентам, що проникають через десмосому з однієї клітини в іншу. 9. Забезпечення скорочувальної функції м'язових волокон. Актинові філаменти є однією з головних частин скорочувального актиноміозинового комплексу. Циклоз. Цитоплазма перебуває в постійному русі, чим забезпечується транспорт речовин до різних ділянок клітини. Рух цитоплазми - універсальній параметр всіх живих клітин. Він забезпечує структурну організацію живої клітини, її енергетичний обмін, веде до збалансованого розповсюдження попередників біосинтезу, продуктів біогенезу, впливає на мембранні потоки. Рух цитоплазми відбувається шляхом коливання, струменевого спрямування, колового руху, фонтануючого руху та ін. Органели цитоплазми • мембранні • немембранні • призначення • принципи функціонування Клітинні органели - диференційовані ділянки цитоплазми, що мають специфічний молекулярний склад. Це складні, високовпорядковані біологічні системи макромолекул, що утворюють певну просторову структуру, здатні до виконання спеціальних клітинних функцій. Клітини тварин містять багато внутрішньоклітинних мембран. Тому майже половина всього об'єму клітин укладена в окремі внутрішньоклітинні відсіки (компартменти), що називаються "органелами" (рис. 1. 29). Інший внутрішньоклітинний простір зайнятий цитозолем. Класифікація органел. Клітинні органели умовно поділяють на мембранні, що оточені типовою біомембраною, і немембранні, що не мають такої оболонки. Мембранні: 1) ендоплазматична сітка: а) зерниста; б) гладенька; 2) комплекс Гольджі; 3) лізосоми; 4) пероксисоми; 5) вакуолі; 6) мітохондрії; 7) пластиди (тільки в рослинних клітинах).
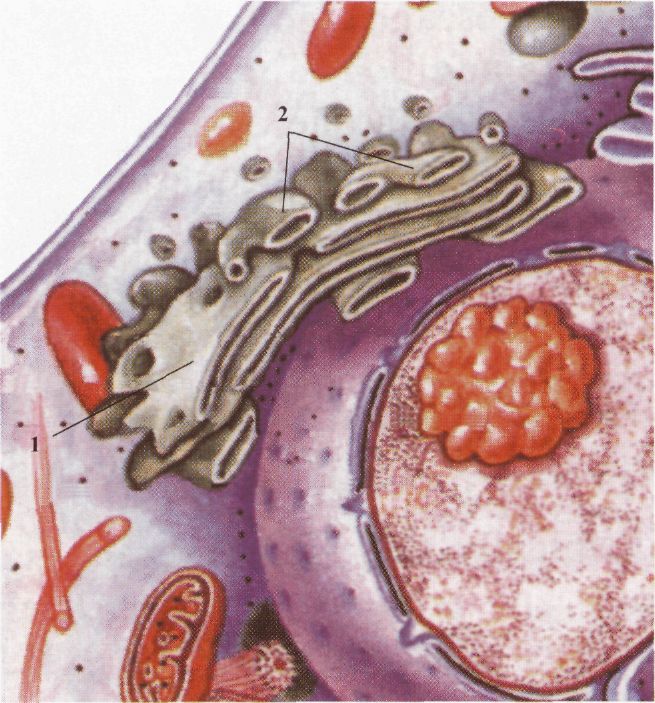
Клітина еукаріотів: / - ядро; 2 - гладенька ендоплазматична сітка; З - зерниста ендоплазматична сітка; 4 рибосоми. Немембранні: 1) рибосоми; 2) центріолі; 3) мікротрубочки; 4) мікрофіламенти. Відповідно до виконуваних функцій розрізняють органели загального і спеціального призначення. Органели загального призначення зустрічаються у всіх еукаріотичних клітинах і належать до загальних структур. Спеціальні органели характерні тільки для певного виду клітин, що виконують специфічну функцію. Наприклад, у деяких найпростіших - це джгутики, скоротлива вакуоля, ундулююча мембрана. У м'язових клітинах — скоротливе волокно; нейрони мають довгі відростки, сперматозоїд - акросому тощо. Ендоплазматична сітка (ЕПС). ЕПС виявлена у всіх еукаріотичних клітинах, відсутня тільки в прокаріотів, у сперматозоїдах і зрілих еритроцитах. ЕПС утворена сіткою мембранних трубочок, цистерн і овальних везикул. ЕПС структурно зв'язана з оболонкою ядра. Розрізняють два типи ЕПС: гладеньку і зернисту, хоча вони структурно пов'язані між собою. Зерниста ЕПС на своїй поверхні містить рибосоми, котрих немає на поверхні гладенької ЕПС. ЕПС утворює сітку мембранних каналів, що пронизують цитоплазму. Ендоплазматична сітка має значення в процесах внутрішньоклітинного обміну, оскільки збільшуєплощу внутрішніх поверхонь клітини, поділяє її на відсіки, що відрізня-
Комплекс Гольджі: 1 - зона формування; 2 - зона дозрівання. ються за фізичним станом і хімічним складом, забезпечує ізоляцію ферментних систем, що, у свою чергу, необхідне для послідовного вступу в узгоджені реакції. Безпосереднім продовженням ендоплазматичної сітки є ядерна мембрана, що відмежовує ядро від цитоплазми, так і зовнішня мембрана (плазмолема), розташована на периферії клітини. Мембранні системи дуже лабільні і можуть змінюватися у залежності від фізіологічного стану клітини, характеру обміну, при рості та диференціюванні. Цитоплазма еукаріотичних клітин містить мембранні шари, пухирці, трубочки, що відокремлюють у сукупності значний внутрішньоклітинний простір. Мембрани ЕПС утворюють безперервні структури із зовнішньою ядерною мембраною, вони спеціалізуються на синтезі й транспорті ліпідів і мембранних білків. Зерниста ЕПС виглядає як система плоских цистерн, зовнішній бік яких вкритий рибосомами, що синтезують білки. Гладенька ЕПС, трубчастої будови, не має рибосом. Гладенька ЕПС зустрічається у клітинах, що виконують секреторну функцію, м'язових і пігментних клітинах. Зерниста ЕПС добре розвинена у клітинах печінки, підшлункової залози, секреторних клітинах, де утворюється білковий секрет. Загальні функції ЕПС. Взаємозалежна система гладенької та зернистої ЕПС працює узгоджено і виконує ряд загальних інтегральних функцій: 1) мембрани ЕПС відокремлюють свій специфічний вміст від цитозолю, утворюють спеціальний ком-партмент; 2) у матриксі ЕПС відбувається нагромадження, збереження і модифікація синтезованих речовин; 3) ЕПС є важливою складовою системи внутрішньоклітинних мембран, забезпечує транспорт синтезованих речовин по внутрішніх порожнинах або за допомогою везикул у різні ділянки клітин; 4) структура ЕПС утворює велику мембранну поверхню всередині клітини, що важливо для багатьох метаболічних реакцій; 5) мембранна система пронизує всю клітину і виступає в якості "внутрішнього скелету". Комплекс Гольджі. Комплекс Гольджі (КГ), утворений комплексом із десятків сплощених дископодібних мембранних цистерн, мішечків, трубочок і везикул, у значній кількості зустрічається в секреторних клітинах. Внутрішній міжмембранний простір заповнений матриксом, що містить спеціальні ферменти. Електронно-мікроскопічні дослідження дозволили переконатися, що КГ збудований із мембран і нагадує стовпчик з порожніх дисків, накладених один на одного. До його складу входить система трубочок із пухирцями на кінцях. Комплекс Гольджі має дві зони: зону формування, куди надходить синтезований матеріал із ЕПС за допомогою транспортних везикул, і зону дозрівання, де формується секрет і зрілі секреторні мішечки. До зони формування надходять синтезовані в ЕПС речовини, що знаходяться в мембранних везикулах. Вони зливаються з мембраною КГ, і вміст везикули надходить всередину комплексу. Речовини обробляються ферментами, після цього знову упаковуються у везикули і переносяться в зону дозрівання. У зоні дозрівання накопичується "дозрілий секрет", що відокремлюється у вигляді секреторних пухирців. У цьому компартменті утворюються також лізосоми і пероксисоми. Функції комплексу Гольджі: 1) нагромадження і модифікація синтезованих макромолекул; 2) утворення складних секретів і секреторних везикул; 3) синтез і модифікація вуглеводів, утворення гліко-протеїдів; 4) КГ відіграє важливу роль у відновленні цитоплазматичної мембрани шляхом утворення мембранних везикул і наступного злиття з клітинною оболонкою; 5) утворення лізосом; 6) утворення пероксисом. Спеціальні функції комплексу Гольджі: 1) формування акросоми сперматозоїда під час сперматогенезу; 2) вітелогенез - процес синтезу і формування жовтка в яйцеклітині. Таким чином, КГ є головним регулятором руху макромолекул у клітині, він збирає синтезовані білки, жири, вуглеводи, формує транспортні везикули і розподіляє по клітині та за її межі. Лізосоми. Лізосоми - це невеликі (0, 2-0, 8 мкм), вкриті мембраною, круглі тільця. Зустрічаються вони у всіх клітинах рослин і тварин, можуть локалізуватися в будь-якому місці клітини. Вміст лізосом складають різні класи гідролітичних ферментів, наприклад, протеази, нуклеази, ліпази, фосфоліпази та ін. Всього нараховується до 40 різних ферментів. Ці ферменти руйнують великі молекули складних органічних сполук, що надходять до клітини (білки, нуклеїнові кислоти, полісахариди). У лізосомах зазнають руйнації мікроорганізми і віруси. Ферменти лізосом перетравлюють зруйновані структури або цілі клітини. Ці процеси називаються аутофагією (від грец. сштос; - самий, фауос; - пожирання). Лізосоми відіграють також істотну роль в індивідуальному розвитку організмів. Вони руйнують тимчасові органи ембріонів і личинок, наприклад, зябра і хвіст у пуголовків жаби, перетинки між пальцями в ембріона людини та ін. Кожна лізосома вкрита щільною мембраною, що ізолює ферменти від цитоплазми. Ушкодження мембран лізосом і вихід із них у цитоплазму ферментів викликає швидке розчинення (лізис) клітини. Втрата лізосомами будь-якої ферментативної системи призводить до тяжких патологічних станів цілого організму, до спадкових хвороб. Вони одержали назву хвороб нагромадження, оскільки пов'язані з нагромадженням у клітинах "неперетравлених" речовин, що заважає нормальному функціонуванню клітини. Ці хвороби можуть виявлятися недостатнім розвитком скелета, окремих внутрішніхсферментів пов'язують розвиток атеросклерозу, ожиріння й інших.
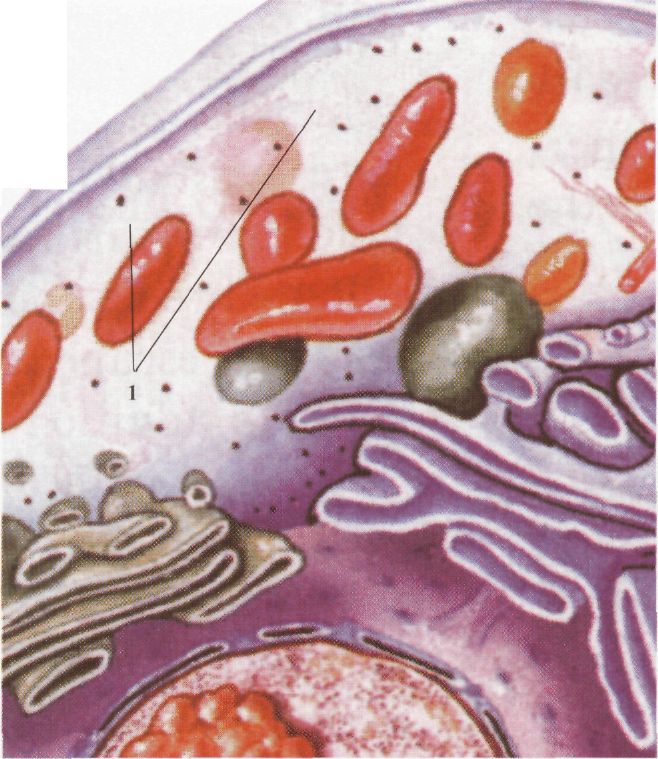
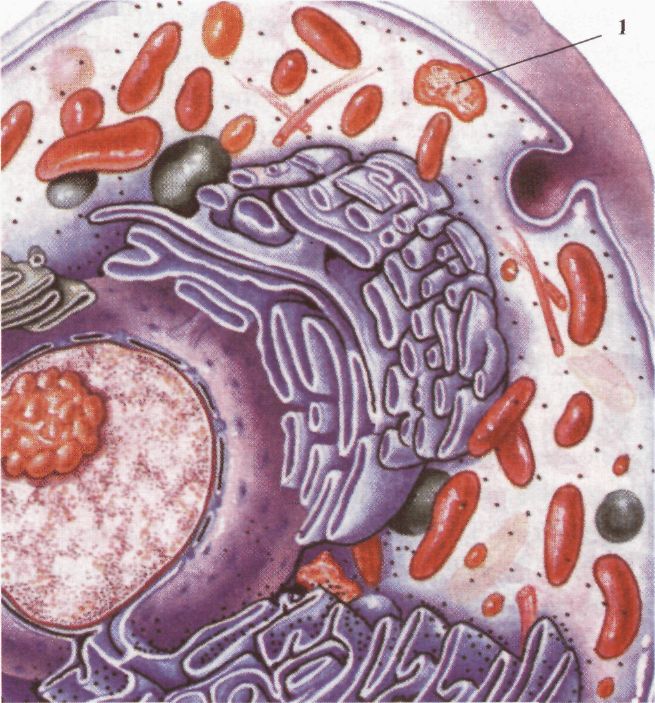
Клітина еукаріотів: 1 - лізосома. З іншого боку, патологічна активність лізосом може спричинити руйнування життєво важливих структур. Лізосоми різноманітні за своєю природою і можуть утворюватися різними шляхами. У кожному випадку формуються морфологічно різноманітні лізосоми, що розщеплюють матеріал із різних джерел. У центрі цих шляхів знаходиться "проміжний компартмент" - ендолізосома. Процес перетравлення лізосомними ферментами об'єктів, що надходять до клітини шляхом фагоцитозу, відбувається у вакуолях, які називаються фа-госомами. Продукти перетравлення потрапляють у цитоплазму, а неперетравлений матеріал залишається у фагосомах і зменшується в розмірах. Такі структури називаються залишковими тільцями. Вони можуть бути різної щільності та розміру. Ендосоми лізосом можуть зливатися з внутрішніми структурами і руйнувати їх. У клітині при цьому утворюються великі мішечки, вкриті спільною мембраною, різної форми і щільності. Такі тільця називаються аутофагосомами. Функції лізосом: 1) перетравлення речовин, що надходять до клітини з навколишнього простору (фагоцитоз), зокрема, таким способом організм бореться з мікробами і вірусами; 2) перетравлення внутрішньоклітинних макромолекул, що виконали свою функцію, і органел (аутофагоцитоз); 3) перетравлення загиблих клітин, або тих, що виконали свою функцію; 4) рециклізація органічних молекул -розщеплення використаних білків, а також вуглеводів, нуклеїнових кислот до мономерів (амінокислот, моносахаридів, нуклеотидів) і повторне їх використання клітиною для синтезу нових молекул. Цим досягається економічність (багатократність) використання внутрішніх молекул. Пероксисоми. Пероксисоми - маленькі сферичні тільця, вкриті мембраною. Виявляються майже у всіх клітинах еукаріотів. їх діаметр становить 0, 3-1, 0 мкм, утворюються в комплексі Гольджі. Пероксисоми містять в основному ферменти для руйнації пероксиду водню, Пероксид водню, що утворюється в результаті окиснення деяких органічних речовин, є токсичним для клітини і тому негайно руйнується каталазою пероксисоми: 2Н2О2 Пероксисоми беруть участь у процесі (3-окис-нення жирних кислот. До 50 % жирних кислот руйнуються в пероксисомах. Вони містять також й інші окисні ферменти. Вакуолі. Вакуолі - це порожнини в цитоплазмі, оточені мембраною та заповнені рідиною. В еукаріотичних клітинах є різні типи вакуоль. Вакуолі можуть виникати з пухирців, які відокремлюються від ендоплазматичної сітки, або комплексу Гольджі. Вони заповнені водним розчином органічних і неорганічних сполук, серед них - продуктів обміну або пігментів. Функції вакуоль різноманітні: вони підтримують тургорний тиск, зберігають поживні речовини і накопичують продукти обміну. Скоротливі вакуолі одноклітинних тварин регулюють осмотичний тиск у клітині, беруть участь у виведенні продуктів обміну, а також сприяють надходженню в клітину води. Мітохондр ії. Мітохондрії (від грец. аітос; - нитка, jov6pa; - зернятко) - це органели, в яких енергія хімічних зв'язків органічних речовин перетворюється на енергію фосфатних зв'язків АТФ. Мітохондрії-досить великі овальні органели
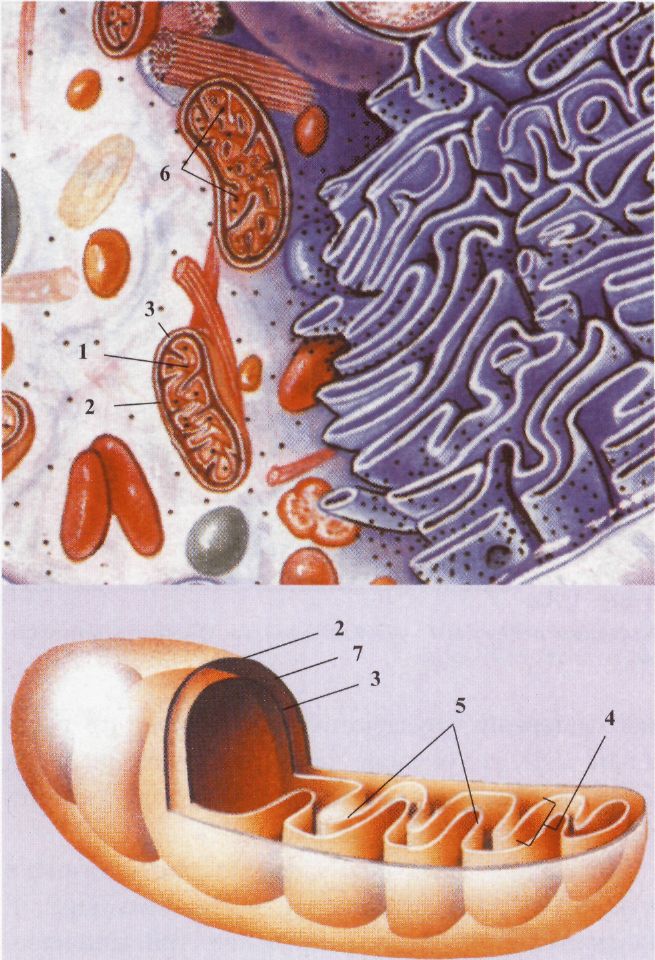
Клітина еукаріотів: / - пероксисома, 2 - вакуолі. рюється на енергію фосфатних зв'язків АТФ. Мітохондрії-досить великі овальні органели (0, 2-2, 0 мкм), вкриті двома мембранами. Вони зустрічаються майже в усіх еукаріотичних клітинах, за винятком анаеробних найпростіших і еритроцитів. Мітохондрії хаотично розподілені по цитоплазмі, хоча частіше виявляються біля ядра або в місцях із високими потребами енергії. У м'язових клітинах вони розташовані між міофібрилами. Органели можуть змінювати свою структуру і форму, здатні переміщуватися всередині клітини. Кількість мітохондрій може змінюватися залежно від активності клітини від кількох десятків до кількох тисяч. Органела містить зовнішню і внутрішню мембрани з вузьким міжмембранним простором. Внутрішня мембрана утворює численні вирости -кристи, що оточені матриксом, в якому знаходиться багато ферментів, рибосоми, одна молекула ДНК. Мітохондрії - органели розміром з бактерію, що використовують енергію окиснення для утворення АТФ. Зовнішня мембрана легко проникна для багатьох невеликих молекул. Містить ферменти, що перетворюють речовини на реакційноздатні субстрати, бере участь в утворенні міжмембранного простору. Внутрішня мембрана погано проникна для більшості речовин. Вона утворює вирости - кристи всередині матриксу. Ця мембрана містить ферменти, що беруть участь у наступних важливих процесах: а) ферменти, що каталізують окисно-відновні реакції дихального ланцюга і транспорту електронів. В результаті утворюється надлишок Н+ у міжмембранному просторі; б) специфічні транспортні білки беруть участь в утворенні градієнту Н+; в) ферментний комплекс АТФ-синтетази, що синтезує АТФ. Міжмембранний простір використовується для градієнта іонів Н+ на внутрішній мембрані, що є необхідною умовою синтезу АТФ. Матрикс - це простір мітохондрії, обмежений внутрішньою мембраною. Він утворений сотнями різних ферментів, що беруть участь у руйнації органічних речовин до CO, і Н2О. При цьому вивільняється енергія хімічних зв'язків між атомами молекул органічних речовин і перетворюється в макроергічні зв'язки АТФ. У матриксі знаходяться рибосоми і молекула мітохондріальної ДНК. Рибосоми міто-хондрій і ДНК забезпечують синтез необхідних орга-нелі білків. Основна інтегральна функція мітохондрій - перетворення енергії та утворення АТФ, що містить макроергічні зв'язки. До мітохондрії надходять різні органічні речовини, які в матриксі окиснюються до найпростіших сполук (СО і НО). Внутрішні мембрани мають необхідний набір ферментів для перетворення енергії хімічних зв'язків, що вивільняється, в енергію АТФ. Піруват і жирні кислоти надходять до мітохондрії і метаболізуються в циклі лимонної кислоти, в якому утворюється НАДН. Потім у процесі окисного фосфорилування багаті на енергію електрони НАДН передаються на кісень за допомогою дихального ланцюга, що зн іходиться на внутрішній мембрані. При цьому в м жмембранному просторі виникає висока концентр; ція Н+. Це створює електрохімічний потенціал на внутрішній мембрані. Завдяки такому хеміосмотичному механізму утворюється АТФ. АТФ-синтетаза, що використовує енергію градієнта Н+ із АДФ та неорганічного фосфату, синтезує АТФ. АДФ + Фн + енергія градієнта Н+ —> АТФ. Енергія АТФ використовується клітиною на рух, скорочення, синтез, транспорт тощо.
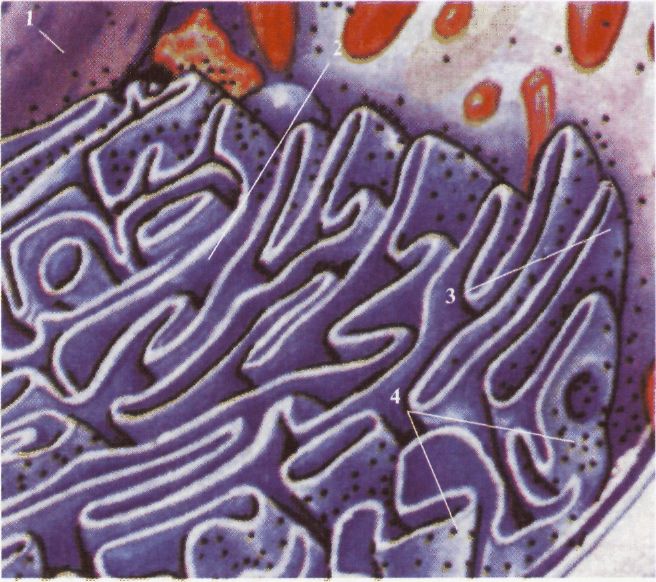
Клітина еукаріотів: 1 - мітохондрія; 2 - зовнішня мембрана; 3 внутрішня мембрана; 4 - криста; 5 - матрикс; 6 - рибосома; 7 - міжміжмембранний простір. Мітохондрії розмножуються шляхом поділу. При поділі клітини вони більш-менш рівномірно розподіляються між дочірніми клітинами. Таким чином між мітохондріями послідовних генерацій клітин здійснюється спадкоємність. Особливості мітохондрій, що вказують на їхню подібність із прокаріотами, розглядають як доказ симбіотичного походження цієї органели. Згідно з такою гіпотезою, деякі аеробні прокаріоти проникли в більшу анаеробну клітину. Можливо, сп
|
||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-09-13; просмотров: 735; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.15 (0.019 с.) |