Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Клітинні мембрани, принцип компартментаціїСодержание книги Поиск на нашем сайте
Основними компонентами клітин є біомембрани, цитоплазм та ядро. Організація і функціонування всіх компонентів клітини пов'язані в першу чергу з біологічними мембранами. Сучасна цитологія розглядає біомембрани як один з основних компонентів клітинної організації, як основу структури і функцій всіх органів і тканин. Більшість клітинних органел мають у своїй основі мембранні структури. Вони характерні для ендоплазматичної сітки, пластинчастого комплексу Гольджі, оболонок і крист мітохондрій, лізосом, вакуоль, пластид, ядерної оболонки і зовнішньої клітинної мембрани. Мембрани - високовпорядковані, складні молекулярні системи, відповідальні за основні процеси життєдіяльності клітин. Наприклад, мембрани поділяють вміст клітини на відсіки (компартменти), завдяки чому в клітині одночасно можуть перебігати різні, навіть антагоністичні, процеси; регулюють метаболічні потоки; підтримують різницю концентрацій речовин (іонів, метаболітів) шляхом переміщення; створюють різницю електричних потенціалів; беруть участь у процесах синтезу і каталізу та ін. Крім того, мембрани є основою для точного розміщення ферментів, а тому зумовлюють впорядкованість обмінних реакцій. Так, в ендоплазматичній сітці відбувається синтез білків, жирних кислот і фосфоліпідів. У мітохондріях здійснюється цикл Кребса, окисне фосфорилування, окиснювання жирних кислот. Існує кілька типів мембран, які відрізняються за будовою, ферментативними властивостями білків, містять різні ліпіди. Так, мембрани мітохондрій тонкі (близько 5 нм) і мають глобулярну структуру білків і специфічний набір фосфоліпідів. Мембрани комплексу Гольджі досить товсті (6-9 нм), містять інші білки і ліпідні молекули. У цитоплазматичних мембранах знаходяться молекули-рецептори до біологічно активних сполук, наприклад, гормонів. Більшість захворювань людини і тварин пов'язані з порушеннями будови і функції мембран. Структура і властивості біомембран. Відповідно до рідинно-мозаїчної моделі будови, клітинні мембрани - це напівпроникний ліпідний бішар із вбудованими в нього білками. Мембрани різних органел мають неоднаковий ліпідний і білковий склад, що забезпечує їх функції. Кожний різновид мембран містить близько 50 % білків. Мембрани мають також значний відсоток вуглеводів. Наприклад, мембрана еритроцитів складається з 40 % ліпідів, 52 % білків і 8 % вуглеводів. Білки не утворюють шари, а розташовані нерівномірно у вигляді мозаїки з глобул; при цьому одні з них знаходяться тільки на поверхні, інші занурені в ліпідну фазу частково або повністю, іноді пронизують її наскрізь.
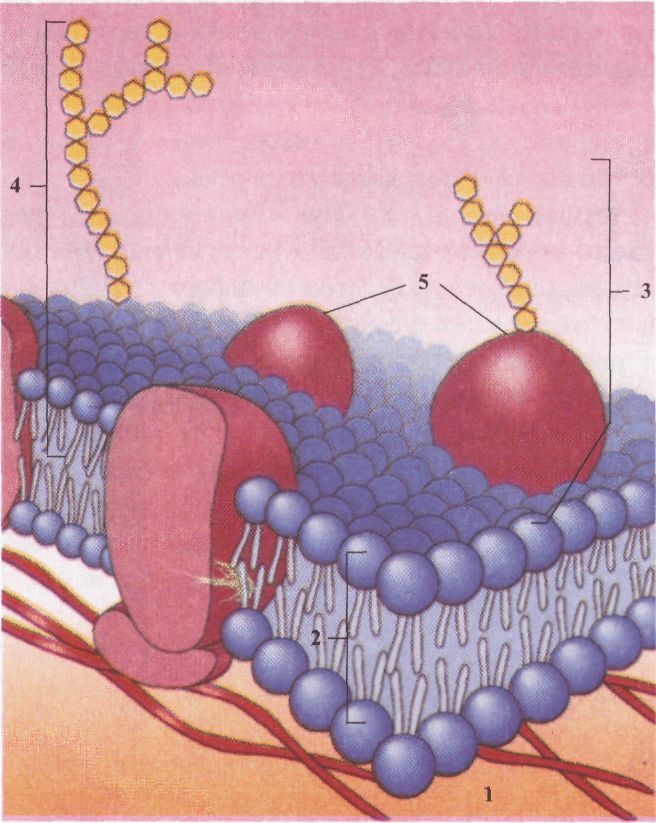
Будова клітинної мембрани: 1 - цитоплазма; 2 - ліпідний бішар; З - глікопротеїн; 4 - гліколіпід; 5 - протеїни.
Ліпідний бішар являє собою рідину, в котрій окремі молекули ліпідів здатні дифундувати в межах свого моношару, але можуть іноді переміщатися з одного шару в другий. В'язкість і рухливість ліпідного бішару залежить від його складу і температури. Цитоплазматична мембрана зовні вкриває клітину і є важливою ланкою в системі біомембран, необхідною умовою існування будь-якої клітини. її поява була однією з умов виникнення життя. Цитоплазматична мембрана має той самий принцип будови, як і інші мембрани. Однак її будова є більш складною, тому що вона є поліфункціональною системою і виконує багато загальних, важливих для всієї клітини функцій. До складу цитоплазматичних мембран, крім ліпідів і білків, входять також молекули гліколіпідів і глікопро-теїдів із розгалуженими вуглеводними ланцюгами. Ці розгалужені ланцюги на поверхні клітини переплітаються один з одним, створюють ніби каркас із вплетеними в нього молекулами білків (глікокалікс), що складається з полісахаридів, ковалентно зв'язаних із глікопротеїдами і гліколіпідами плазмолеми. Функції глікокаліксу: а) міжклітинне розпізнавання; б) міжклітинна взаємодія; в) пристінкове травлення. З внутрішнього боку клітини білки і глікопротеї-ди зв'язані з мікротрубочками і білковими фібрила ми, що складають елементи цитоскелета. Часто плазматична мембрана утворює безліч пальцеподібних виступів — мікроворсинок. Це значно збільшує всмоктувальну поверхню клітин, полегшує перенесення речовин через зовнішню мембрану та їх прикріплення до поверхні субстрату. Ліпіди біомембран. Мембранні ліпіди - амфі-патичні молекули (володіють як гідрофобними, так і полярними властивостями) і у водному середовищі утворюють подвійний шар (бішар). Ці бішари са-моорганізуються у закриті компартменти, що здатні відновлюватися при ушкодженнях. Розрізняють три основних класи ліпідних молекул - фосфоліпіди, холестерин і гліколіпіди. За складом внутрішній і зовнішній шари мембран відрізняються один від одного. Різний ліпідний склад характерний як для всіх типів клітин, так і для різних органел однієї і тієї ж еукаріотичної клітини. Ліпідний бішар є розчинником для мембранних білків, які функціонують тільки в присутності певних ліпідів. Ліпідний бішар мембран асиметричний, що забезпечує правильну орієнтацію білків, і має напівпроникні властивості. Білки біомембран. Білки складають понад 50 % від маси мембран, більшість із них має глобулярну структуру. Частина мембранних білків можуть вільно переміщуватися у фосфоліпідному бішарі, але здебільшого фіксовані в певних місцях у площині мембран. Мембранні білки розподілені по зовнішньому і внутрішньому бішарах нерівномірно (асиметрично). Для мембран різних органел характерний неоднаковий білковий склад. Групи білків мембрани, які розташовані в одному місці і зв'язані один з одним, утворюють групи (кластери), що виконують загальну функцію, наприклад, транспорт електронів у дихальному ланцюгу мітохондрій. Деякі мембранні білки фіксовані в бішарі мікрофіламен-тами і мікротрубочками цитоскелета. Ліпідний бішар визначає основні структурні особливості біологічних мембран, тоді як білки відповідальні за більшість мембранних функцій. Функції біологічних мембран. Мембрани беруть участь у виконанні різноманітних функцій. Причому функції біомембран у значній мірі визначають властивості і фізіологію клітини. Наприклад, секреторні клітини містять багато мембран апарату Гольджі та ендоплазматичної сітки. Нервові клітини мають мембранні відростки (дендрити й аксони), що проводять електричні імпульси. М'язові клітини містять дуже багато мітохондрій. Мембрана складається з ділянок (кластерів), що мають свій набір ліпідів, білків та інших молекул. Специфічність комплексного набору макромолекул визначає функціональну особливість ділянки мембрани. Внаслідок цього на різних ділянках мембрани можуть одночасно проходити різні процеси. Наприклад, на внутрішній мітохондріальній мембрані відразу відбувається декілька процесів, що точно скоординовані та є частинами однієї інтегральної функції - перетворення енергії. Транспорт реч овин через мембрану. Ліпідний бішар практично непроникний для більшості полярних водорозчинних молекул, оскільки внутрішня частина його гідрофобна. Завдяки такому бар'єру запобігається втрата водорозчинного вмісту клітини. Різні речовини мають неоднакову здатність проникати через цей бар'єр. Великі макромолекули (білки, жири) та їх агрегати не можуть проникати крізь мембрану. Для перенесення існує "макромеханізм" - захоплення клітиною і доставка в певному напрямку (ендоци-тоз і екзоцитоз). Невеликі молекули переносяться за допомогою спеціальних молекулярних механізмів через мембрану: шляхом пасивного й активного транспорту. Транспорт невеликих молекул. Є два типи транспорту молекул через мембрану: пасивний і активний. Пасивний транспорт - переміщення невеликих полярних (СО2, Н2О) і неполярних (О2, N2) молекул за градієнтом концентрації або електрохімічним градієнтом без витрати енергії. Існують різні форми пасивного транспорту: 1. Проста дифузія газів при диханні між порожниною альвеол легень і просвітом кровоносних капілярів (аерогематичний бар'єр). Характеризується низькою вибірковістю мембрани до речовин, що переносяться. 2. Полегшена дифузія, за участю компонентів мембрани (канали і переносники) переважно в одному напрямку (у клітину) за градієнтом концентрації без витрат енергії, характеризується вибірковістю до речовин. 3. Осмос - процес дифузії розчинника (Н2О) через напівпроникну мембрану за концентраційним градієнтом із високої концентрації розчинника у бік з низькою концентрацією. Клітина має два класи мембранних транспортних білків, що формують наскрізні шляхи через гідрофобний шар: численні білки-переносники й іонні канали. Білки-переносники - це складні глобулярні білки, що мають спорідненість до певних молекул, забезпечують їх перенесення через мембрану. Іонні канали — складаються із кількох зв'язаних між собою білкових субодиниць, що формують у мембрані велику пору. Через неї за електрохімічним градієнтом проходять іони. Активний транспорт - перенесення молекул через мембрану за допомогою спеціальних білків проти концентраційного або електрохімічного градієнта з використанням енергії АТФ. Білки-переносники є одночасно ферментами і називаються АТФ-азами. Транспорт агрегатів великих молекул. Ендоцитоз - складний активний процес поглинання клітиною великих молекул, часток, мікроорганізмів. Різновиди: піноцитоз, фагоцитоз, опосередкований рецепторами ендоцитоз. Піноцитоз - поглинання рідини та розчинених речовин з утворенням специфічних мембранних пухирців.
Зріз тканини легені: 1 - альвеоли; 2 - кровоносні судини.
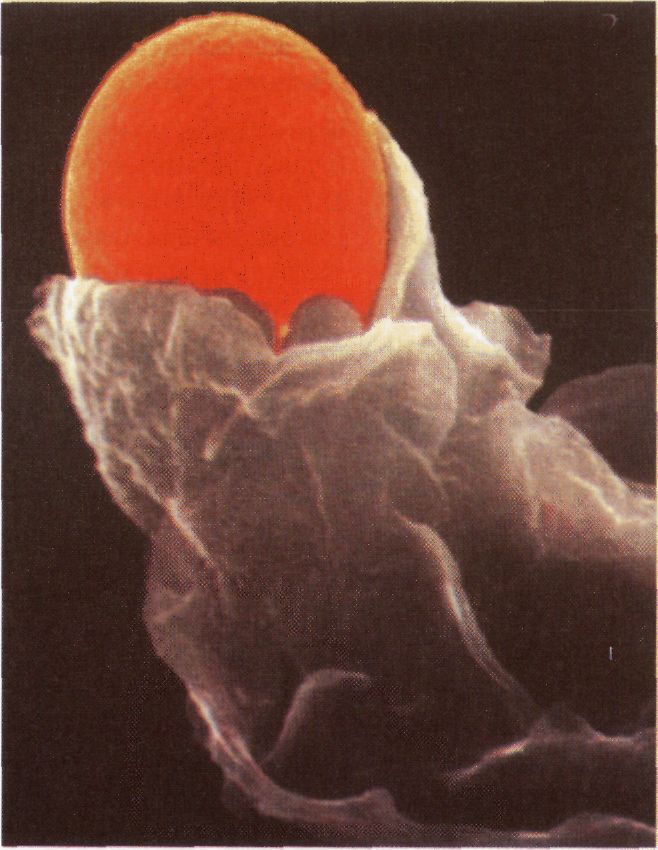
Фагоцитоз - поглинання твердих часток (мікроорганізмів, часток клітин). При цьому утворюються великі щільні ендоцитозні пухирці - фагосоми, які зливаються з лізосомами і формують фаголізосоми. Опосередкований рецепторами ендоцитоз характеризується поглинанням із позаклітинної рідини певних макромолекул. Екзоцитоз - процес виведення макромолекул, при якому внутрішньоклітинні секреторні пухирці зливаються з плазмолемою і їх вміст виводиться з клітини. При ендоцитозі ділянка плазматичної мембрани втягується і замикається: Утворюється ендоцитоз-ний пухирець, що містить поглинений матеріал. При екзоцитозі мембрана секреторних пухирців зливається з плазматичною мембраною і вміст вивільняється в позаклітинний простір. Компартментація клітини. Компартментація - це просторове розмежування клітини внутрішніми мембранами на відсіки, що дозволяє здійснювати перебіг багатьох біохімічних реакцій (часто протилежно спрямованих) одночасно і незалежно одна від одної. Близько половини загального об'єму еукаріо-тичних клітин розділені мембранами на компарт-менти, відомі як клітинні органели. Кожний відсік має у своєму складі специфічні білки, що визначають його унікальні функції. Крім того, цитозоль, ендоплазматичний ретикулум, комплекс Гольджі, ядро, мітохондрія, ендосома, лізосома і пероксисома, -це індивідуальні компартменти, відділені від іншихвідсіків клітини принаймні однією відбірково проникною мембраною.
Фагоцитоз.
Внутрішні мембрани еукаріотичної клітини роблять можливою функціональну спеціалізацію різних частин клітини, що є вирішальним фактором у роз'єднанні безлічі процесів, що одночасно проходять у клітині. Ядро містить основну частину спадкового матеріалу і є головним місцем синтезу ДНК і РНК. Цитоплазма, що оточує ядро, складається з цито-золю та розташованих у ньому цитоплазматичних органел. Об'єм цитозолю становить більше половини від загального об'єму клітини. Саме в ньому синтезуються білки і перебігає більшість метаболічних реакцій, в яких одні молекули руйнуються, а інші утворюються, забезпечують необхідні будівельні блоки. Близько половини всіх мембран клітини обмежують схожі на лабіринт порожнини ендоплазматичного ретикулуму (ЕПР). На зверненому до цитозолю боці ЕПР знаходиться багато рибосом. Рибосоми беруть участь у синтезі різних білків, призначених для секреції або для інших органел В ЕПР також синтезуються ліпіди для клітини. Вважається, що всі цистерни зернистого і гладенького ЕПР зв'язані між собою й утворюють єдиний великий компартмент. Комплекс Гольджі складається з розміщених один над одним сплощених мембранних мішечків, що називаються цистернами Гольджі; він одержує з ЕПР білки, ліпіди і відправляє ці молекули в різні ділянки клітини, піддаючи їх модифікаціям. Мітохондрії і хлоропласти рослинних клітин виробляють велику частину АТФ, що використовується для всіх видів роботи клітини. Лізосоми містять ферменти, що руйнують органели, а також частки і молекули, поглинені клітиною ззовні шляхом ендоцитозу. Нарешті, пероксисоми беруть участь у процесах нейтралізації пероксиду водню та інших токсичних метаболітів. Рецептори клітин Для регуляції поділу, росту, розвитку, організації й обміну інформацією, координації функцій клітини взаємодіють між собою. Це відбувається шляхом виділення хімічних речовин і утворення щілиноподібних контактів. Крім цього, на плазматичних мембранах клітини розташовані сигнальні молекули - білки, які отримали назву рецептори. Рецептори зв'язують молекулу й ініціюють відповідь. Вони представлені трансмембранними білками, що мають спеціальну ділянку для зв'язування фізіологічно активних молекул: гормонів і нейромедіаторів. Багато рецепторних білків у відповідь на зв'язування певних молекул змінюють транспортні властивості мембран. Внаслідок цього може змінюватися полярність мембран, генеруватися нервовий імпульс або змінюватися обмін речовин. Розрізняють внутрішньоклітинні рецептори і рецептори, що розташовуються на поверхні клітини у плазматичній мембрані. Серед них виділяють рецептори двох типів - зв'язані з каналами клітини і не зв'язані з каналами. Вони різняться між собою за швидкістю та вибірковістю впливу сигналу на певні мішені. Рецептори, зв'язані з каналами, після взаємодії з хімічними речовинами (гормон, нейро-медіатор) сприяють утворенню в мембрані відкритого каналу, в результаті чого зразу ж змінюється її проникність. Рецептори, не зв'язані з каналами, також взаємодіють з хімічними речовинами, але іншої природи, здебільшого це ферменти. Тут ефект опосередкований, відносно сповільнений, але більш тривалий. Функція цих рецепторів лежить в основі навчання і пам'яті. Більшість клітин багатоклітинного організму спеціалізована на виконанні однієї головної функції, і всі вони наділені характерним набором рецепторів. Це дозволяє відповідно реагувати на хімічні сигнали, які запускають або модулюють дану функцію. Багато з сигнальних молекул діють в дуже низьких концентраціях (переважно не більше 108М), і рецептори, які з ними зв'язуються, мають також високу спорідненість (константа спорідненості близько 108 л/моль). Одна і та ж сигнальна молекула у клітинах-мішенях може викликати різні ефекти. Так, ацетилхолін стимулює скорочення волокон скелетних м'язів, але водночас зменшує частоту і силу скорочень м'яза серця. Такі різні ефекти зумовлені відмінностями рецепторів. Число рецепторів до певної речовини (ліганда) може коливатися в межах від 500 до 100 000 на клітину і вони розташовуються на мембрані випадково або сконцентровані в певних її ділянках. Рецептори клітинної поверхні складають не більше 0,1 % всієї маси білка плазматичної мембрани і тому їх важко виділити у чистому вигляді. Ця перешкода долається застосуванням методів кло-нування послідовностей ДНК, які кодують поверхневі рецептори.
|
||||
|
|
Последнее изменение этой страницы: 2016-09-13; просмотров: 1203; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.205.110 (0.009 с.) |