Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Мітохондрії, їх будова і функціїСодержание книги
Поиск на нашем сайте
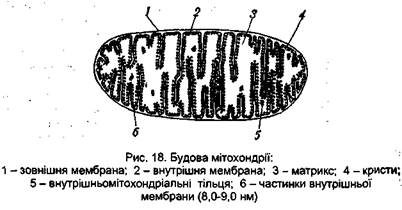
Сукупність реакцій дихального ланцюга і процес окиснювального фосфорилювання відбуваються в мітохондріях – субклітинних органелах специфічної будови. Кількість їх у клітині коливається від декількох сотень до декількох десятків тисяч. Розміри І форма мітохондрій дуже різноманітні і залежать від типу клітин. Вони бувають циліндричної, сферичної або ниткоподібної форми, а також пластинчастої та зірчастої форм. Проте мітохондрії, виділені з різних джерел, мають багато спільного. Оболонка цих органел складається з двох мембран – зовнішньої і внутрішньої. Зовнішня мембрана гладка, вона наче
обтягує і формує мітохондрію. Внутрішня мембрана утворює перпендикулярні до поверхні мембрани численні складки, або виступи, названі кристами, які вдаються в тіло мітохондрій досить далеко, досягаючи навіть протилежної стінки. Завдяки наявності крист внутрішня мембрана має дуже велику довжину. Внутрішній простір мітохондрій заповнено рідиною, яка називається матриксом. Мембрани складаються на 1/3 зліпідів і на 2/3 – з білків, переважно структурних. Близько 25% загального білка внутрішньої мембрани складають білки ферментів, що здійснюють перенесення електронів і процеси фосфорилювання. Цими ферментами є НАД-залежні дегідрогенази, флавопротеїди, цитохроми b, с, а і а3
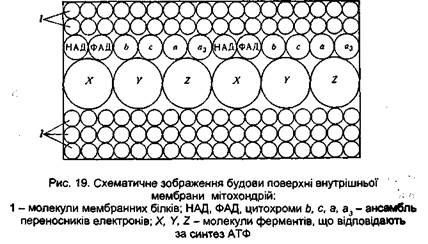
Останнім часом установлено, що ферменти мітохондрій згруповані в компактні агрегати молекулярною масою 1 млн., які називають дихальними ансамблями (рис. 19). Дихальні ансамблі вмонтовані поперек внутрішньої мембрани мітохондрій і рівномірно розподілені в ній. Кількість таких ансамблів у кожної мітохондрії складає 5000-10000. Слід зазначити, що функціональна активність цих ансамблів виявляється тільки в сполученні їх із мембраною мітохондрій. Відділення їх від мембрани призводить до втрати здатності до спряженого фосфорилювання, хоча вони і здатні переносити електрони. Без мембрани окремі ферменти дихального ланцюга стають роз'єднаними, розташовуються хаотично і безсистемно, у результаті чого втрачають здатність утилізувати енергію, яка виділяється при окисненні. Через це вважають, що функції мітохондрій абсолютно унікальні. Оскільки саме в них відбувається переважно синтез макроергічних сполук, мітохондрії називають ще силовими станціями клітини. Не варто думати, що будь-яке окиснення субстратів у живих системах супроводжується фосфорилюванням. Передавання атомів водню й електронів ферментами дихального ланцюга і зв'язане з цим фосфорилювання АДФ здійснюються всередині мітохондрії, на її внутрішній мембрані, де окремі ферменти ланцюга перенесення водню й електронів сполучені в суворо визначеному порядку, створюючи своєрідний комплекс. Саме таке структурне сполучення ферментів із мембраною забезпечує використання енергії окиснення. Процес передачі атомів водню й електронів відбувається і на поверхні мітохондрій, і в гіалоплазмі за участю тих же ферментів, що й окиснення, спряжене з фосфорилюванням. Проте внаслідок того, що ці ферменти не зв'язані мітохондріальною мембраною, вони розміщені хаотично і тому окиснення тут не супроводжується активацією неорганічного фосфату. У цьому випадку енергія окиснення перетворюється в тепло, і такий процес називається вільним окисненням. На даний час установлено, що процеси окиснювального фосфорилювання і вільного окиснення в клітинах і в організмі взаємозв'язані. За певних умов окиснювальне фосфорилювання може переключатися на вільне окиснення. Таке переключення можна спостерігати при охолодженні організму, коли зниження температури призводить до роз'єднання ферментів окиснювального фосфорилювання і воно переходить у вільне окиснення для посилення продукції тепла. На закінчення слід сказати, що спряжене окиснювальне фосфорилювання є основним, але не єдиним способом синтезу АТФ в органах і тканинах тваринного організму. Невелика кількість АТФ утворюється шляхом так званого субстратного фосфори-лювання. У цьому випадку активування неорганічного фосфату відбувається за участю самого субстрату. Тому таке фосфорилювання часто називають фосфорилюванням на рівні субстрату, щоб відрізнити його від реакцій фосфорилювання, що відбуваються за участю дихального ланцюга. При субстратному фосфорилюванні між субстратом і неорганічним фосфатом спочатку утворюється макроергічний зв'язок за рахунок окиснення субстрату. Енергія його окиснення передається залишку фосфатної кислоти. Активований у такий спосіб неорганічний фосфат передається на АДФ з утворенням АТФ. Субстратне фосфорилювання – дуже давній спосіб біосинтезу АТФ. Він виник в організмах, коли на Землі не було кисню, тобто в анаеробних умовах. Проте клітини тваринного організму зберегли здатність синтезувати АТФ шляхом субстратного фосфорилювання і здійснюють його за умови дефіциту кисню в тканинах.
Обмін вуглеводів ТРАВЛЕННЯ ВУГЛЕВОДІВ Вуглеводи займають дуже важливе місце в харчуванні людини і тварин. Вони складають більшу частину (близько 60-70%) харчового раціону людини і є основним джерелом енергії, необхідної організму для здійснення різних функцій. Тому кількість вуглеводів як основного енергетичного матеріалу повинна збільшуватися в раціоні людини в міру посилення фізичного навантаження. Джерелом вуглеводів для людини є продукти рослинного походження - мука, різноманітні крупи, картопля. Основним вуглеводом цих продуктів є крохмаль і в незначній кількості клітковина. З фруктами, ягодами і харчовими цукрами людина одержує переважно сахарозу і фруктозу. Проте не всі вуглеводи засвоюються організмом людини однаково. Клітковина, наприклад, не перетравлюється травними соками. Вона лише частково розщеплюється за допомогою бактерій у товстому відділі кишок до дисахариду целобіози і моносахариду глюкози. Тут же мікроорганізми використовують клітковину для біосинтезу вітамінів групи К. Клітковина необхідна як стимулятор секреторної і моторної функцій травного каналу і для формування калу, тому вона є важливою складовою частиною харчового раціону людини. Проте надмірне надходження клітковини в організм викликає посилення перистальтики кишок, і їжа проходить через травний канал дуже швидко. У результаті цього харчові продукти не повністю перетравлюються і всмоктуються, значна частина їх потрапляє в кал і видаляється з організму невикористаною. Поряд із цим при підвищеній кількості клітковини в харчовому раціоні в товстому відділі кишок посилюються процеси бродіння, відбувається скупчення газів, що може призвести до серйозних ускладнень у хворих людей, а також у тих, хто мало рухається. Такі полісахариди, як крохмаль і глікоген, що надходять із їжею, попередньо підлягають розщепленню в травному каналі під дією певних ферментів. У результаті розщеплення утворюються моносахариди, які потім легко асимілюються тканинами організму. Процес перетравлювання вуглеводів (крохмалю і дисахаридів) починається вже в ротовій порожнині під дією двох ферментів, що містяться в складі слини, - амілази і а-глюкозидази (мальтази). Амілаза розщеплює крохмаль, а-глюкозидаза - мальтозу. Амілаза слини є більш активною порівняно з мальтазою, тому в ротовій порожнині з крохмалю утворюються переважно великі фрагменти його молекул - декстрини, що мають солодкий смак, і лише незначна кількість мальтози і глюкози. Оскільки в ротовій порожнині їжа довго не затримується, подальше розщеплення вуглеводів продовжується в наступних відділах травного каналу. У шлунку немає ферментів, які розщеплюють вуглеводи. До того ж ферменти слини припиняють свою дію в шлунку, де реакція середовища кисла, а оптимум дії цих ферментів лежить у нейтральному або слабокислому середовищі. У дванадцятипалій кишці й у порожнині тонкої кишки під дією ряду ферментів підшлункової залози і кишкового соку відбувається остаточне розщеплення вуглеводів до моносахаридів. Так, а-амілаза розщеплює крохмаль і декстрини до мальтози, α-глюкозидаза розщеплює мальтозу на дві молекули глюкози. Під дією β-галактозидази (лактази) кишкового соку лактоза розщеплюється на глюкозу і галактозу, β-фруктофуранозидаза (сахараза) розщеплює сахарозу на глюкозу і фруктозу. Моносахариди, що утворилися, глюкоза, фруктоза, галактоза добре розчинні у воді - є тими простими цукрами, що всмоктуються стінкою кишок і через капіляри кишкових ворсинок потрапляють у кров'яне русло. Під час усмоктування частина моносахаридів (фруктоза, галактоза і маноза) перетворюються в глюкозу. У такий спосіб із кишок у кров потрапляє переважно глюкоза, і лише в незначних кількостях - фруктоза, галактоза і маноза. З током крові через ворітну вену вони потрапляють у печінку й інші органи і тканини. Частина глюкози затримується в печінці і використовується переважно для біосинтезу глікогену й інших складних вуглеводів, частина перетворюється в жири й окиснюється в тканинах. За нормальних умов на синтез глікогену використовується 3-5% глюкози, 30-35% її перетворюється в жири, а основна маса (60-70%) окиснюється до СО2 і H2О з виділенням енергії. Невеличка кількість глюкози є обов'язковою складовою частиною крові. За нормою це складає до 120 мг%. Цей рівень підтримується, незважаючи на постійне споживання глюкози тканинами і періодичне надходження її" з кишок після прийому їжі. Головна роль у підтримці постійної концентрації глюкози в крові належить печінці. За недостатньої кількості глюкози в крові під впливом нервових імпульсів, що йдуть із центральної нервової системи, частина глікогену печінки розщеплюється, підвищуючи тим самим рівень глюкози в крові. Цей процес називають мобілізацією глікогену. Регуляторну роль у підтримці постійного рівня глюкози в крові центральна нервова система здійснює не тільки шляхом прямого впливу на печінку, але й через ендокринні залози, серед яких найбільше значення мають надниркові і підшлункова залози. У мозковій речовині надниркових залоз утворюється адреналін, підвищений вміст якого в крові стимулює розщеплення глікогену в печінці. Підшлункова залоза виробляє гормон інсулін. Підвищене виділення цього гормону в кров супроводжується зниженням глюкози в крові. Таким чином, інсулін діє протилежно адреналіну: він затримує розпад глікогену в печінці і сприяє відкладенню його в м'язах. Крім інсуліну, в підшлунковій залозі виробляється ще один гормон - глюкагон, дія якого протилежна інсуліну. Істотний вплив на рівень глюкози в крові має і ряд інших гормонів: соматотропін, кортикостерон, тироксин. Всі ці гормони викликають підвищення вмісту глюкози в крові, через що їх називають діабетогенними (від назви захворювання "діабет", характерним для якого є підвищений вміст цукру в крові в результаті порушення функції підшлункової залози, що виробляє інсулін). Концентрація глюкози в крові може збільшуватися до 160-180 мг% при вживанні з їжею великої кількості вуглеводів. Це явище зветься аліментарною, або харчовою, гіперглікемією, що супроводжується виділенням глюкози із сечею, тобто глюкозурією. З вищевикладеного випливає, що надходження глюкози в кров відбувається переважно двома шляхами: розщепленням глікогену в печінці й усмоктуванням простих цукрів із кишок після прийому їжі. Утворення глюкози з глікогену відбувається не прямим його розщепленням, а за допомогою ферментів шляхом приєднання до нього фосфатної кислоти. При цьому від глікогену послідовно відщеплюються залишки глюкози у вигляді фосфорних ефірів, які потім розщеплюються на глюкозу і фосфатну кислоту:
Невелика кількість глюкози утворюється також шляхом гідролітичного розщеплення глікогену, проте цей шлях має дргорядне значення.
|
||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 759; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.89.197 (0.007 с.) |