Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тема: обмін речовин і енергіїСтр 1 из 11Следующая ⇒
Тема: Біологічне окиснення
Історія розвитку уявлення про біологічне окиснення Питання про те, яким чином відбувається окиснення речовин в організмі, давно привертало увагу вчених. Ще А. Лавуазьє виявив, що між процесом горіння органічних речовин поза організмом і процесом дихання тварин є багато спільного. Виявилося, що при диханні, як і при горінні, із повітря поглинається кисень і утворюються СО2 і Н2О, тобто в організмі відбувається процес, подібний горінню або окисненню. Проте було незрозуміло, чому горіння речовин в організмі відбувається за низької температури (37° С), без полум'я і до того ж у водному середовищі, адже за звичайних умов вода припиняє горіння. Тому було запропоновано ряд теорій окиснення речовин в організмі. Однією з перших була теорія К. Шенбайна. Він відкрив озон, здатний окиснювати деякі речовини за звичайної температури. Оскільки озон являє собою "активний кисень", К. Шенбайн зробив припущення про те, що кисень, який вдихається з повітрям, в організмі активізується. У подальшому ця теорія була розвинута російським ученим О.М. Бахом. Згідно з теорією О.М. Баха, активація кисню в організмі відбувається за допомогою речовин, які легко окиснюються, у результаті чого утворюються пероксиди. Такими речовинами, що легко окиснюються, є оксигенази. Далі атом кисню перекису переноситься на інші речовини, які важче окиснюються за допомогою ферменту пероксидази. Ця теорія схематично виглядає так:
Оскільки шлях кисню до речовини, що окиснюеться, лежить через пероксиди, теорія О.М. Баха одержала назву пероксидної теорії окиснення. Вивчаючи окиснення речовин у рослинах, російський біохімік В.І. Палладій установив, що воно може відбуватися й за відсутності кисню, якщо в реакційному середовищі є речовини, здатні приєднувати атоми водню. Такими речовинами в рослинах є пігменти, або так звані хромогени, які й були виявлені В.І. Палладіним. Приєднуючи атоми водню від субстратів, що окиснюються, тобто відновлюючись, вони стають безбарвними, а віддаючи його, або окиснюючись, вони забарвлюються і перетворюються в пігменти. У безбарвні хромогени ці пігменти перетворюються за наявності кисню. На думку В.І. Палладіна, кисень повітря не досягає субстрату і безпосередньо з ним не взаємодіє, роль його полягає тільки в окисненні атомів водню хромогенів з утворенням молекул води. Різні речовини, що окиснюються в організмі, відщеплюють водень за допомогою ферментів дегідрогеназ. Таким чином, відбувається процес дегідрування, що і с процесом окиснення речовин. У своїй теорії В.І. Палладій надавав великого значення кисню як акцептору водню, розкривши тим самим важливу роль кисню в біологічному окисненні.
Доказом справедливості теорії В.І. Палладіна стали праці Г.О. Віланда, який показав, що окиснення спиртів і альдегідів також можливе за відсутності кисню І відбувається шляхом дегідрування. На підставі цього Г.О. Віланд висловив припущення про те, що дегідрування субстратів і є головним процесом, що лежить в основі біологічного окиснення, і що кисень реагує безпосередньо з активованими атомами водню. Теорія окиснення речовин шляхом дегідрування, або "активації", водню субстратів, що окиснюються, називається теорією Палладіна-Віланда. Надалі підтвердженням справедливості даної теорії стало відкриття в 1900-1920 pp. цілого ряду дегідрогеназ – ферментів, що віднімають водень від субстратів, які окиснюються. У 1913 р. О.Г. Варбург виявив, що невеликі кількості ціаніду майже цілком припиняють поглинання кисню клітинами і тканинами, що дихають. Оскільки ціаніди утворюють стійкі комплекси з залізом, О.Г. Варбург припустив, що в процесі окиснення речовин велику роль відіграє якийсь універсальний залізовмісний фермент, здатний переносити електрони й активувати кисень. О.Г. Варбург назвав його дихальним ферментом. Дещо пізніше Д. Кейлін довів фундаментальну роль ще ряду залізовмісних ферментів, які називаються цитохромами і дуже схожі на дихальний фермент Варбурга. Д. Кейлін, поєднавши дві існуючі теорії окиснення речовин (приєднання кисню і дегідрування), показав, що процеси окиснення ряду субстратів включають як дегідрування, так і окиснення киснем. Процес окиснення субстратів являє собою ланцюг послідовних реакцій, які починаються реакціями відщеплення водню і закінчуються перенесенням електронів на кисень. Через те, що при такому окисненні відбувається постійне поглинання кисню, цей процес окиснення субстратів називається тканинним диханням.
Викладені вище теорії (теорія Шєнбайна - Баха про активацію кисню і теорія Палладіна - Віланда про активацію субстратів, тобто водню) склали основу сучасного уявлення про механізм біологічного окиснення. Окиснювальне фосфорилювання Для різних життєвих процесів організму необхідна енергія. Під час біологічного окиснення, або тканинного дихання, яке ми розглянули вище, відбувається виділення вільної енергії, яка використовується в організмі у двох напрямках: частина її споживається для різних реакцій біосинтезу, частина використовується для підтримки постійної температури тіла, тобто перетворюється в тепло. Для того щоб вільна енергія окиснення субстратів могла бути використана для реакцій синтезу, скорочення м'язів і т.д., необхідно, щоб вона набула доступної для цього форми. На початку 1930-х років академік В.О. Енгельгардт висловив ідею про те, що при кисневому диханні відбувається фосфорилювання аденозиндифосфатної кислоти (АДФ) з утворенням аде-нозинтрифосфатної кислоти (АТФ). Пізніше, у 1939-1940 pp., радянський біохімік В.О. Беліцер установив, що при окисненні різних субстратів, зокрема, янтарної і лимонної кислот, відбувається споживання неорганічної фосфатної кислоти й утворення АТФ. Аденозинтрифосфатна кислота являє собою універсальне рухливе джерело хімічної енергії в клітинах. Ця енергія зосереджується, або акумулюється, у пірофосфатних хімічних зв'язках молекул АТФ:
Кількість енергії, акумульованої в одному макроергічному зв'язку АТФ, складає 33,5-41,9 кДж. При гідролізі АТФ хімічна енергія макроергічних зв'язків звільняється і може бути використана для різних життєвих процесів. Таким чином, вільна енергія окиснення субстратів перетворюється в доступну для організму хімічну форму, акумульовану в пірофосфатних зв'язках АТФ, молекули якої є формою накопичення цієї енергії. Для здійснення процесу фосфорилювання, тобто з'єднання АДФ з неорганічною фосфатною кислотою, неорганічний фосфат має бути активований. Джерелом енергії для його активації служить енергія окиснення субстратів. Таким чином, процес фосфорилювання АДФ, що супроводжується утворенням АТФ, пов'язаний з процесом окиснення, тому називається окиснювальним фосфорилюванням. Саме таким шляхом в організмі синтезується значна кількість АТФ. У цьому процесі бере участь дихальний ланцюг ферментів, за допомогою яких атоми водню й електрони, відняті від субстратів, передаються на кисень. Сам же субстрат у цьому процесі участі не бере. Як показав В.О. Беліцер, при перенесенні однієї пари електронів (або двох атомів водню) від субстрату до кисню утворюється три молекули АТФ. На даний час відомо, в яких місцях дихального ланцюга відбувається активування неорганічного фосфату і фосфорилювання АДФ. Перше активування відбувається на ділянці між НАД і флавопротеїдом, друге - між цитохромами b и с, і трете – між цитохромом а3 і молекулярним киснем: < і Обмін вуглеводів ТРАВЛЕННЯ ВУГЛЕВОДІВ Вуглеводи займають дуже важливе місце в харчуванні людини і тварин. Вони складають більшу частину (близько 60-70%) харчового раціону людини і є основним джерелом енергії, необхідної організму для здійснення різних функцій. Тому кількість вуглеводів як основного енергетичного матеріалу повинна збільшуватися в раціоні людини в міру посилення фізичного навантаження.
Джерелом вуглеводів для людини є продукти рослинного походження - мука, різноманітні крупи, картопля. Основним вуглеводом цих продуктів є крохмаль і в незначній кількості клітковина. З фруктами, ягодами і харчовими цукрами людина одержує переважно сахарозу і фруктозу. Проте не всі вуглеводи засвоюються організмом людини однаково. Клітковина, наприклад, не перетравлюється травними соками. Вона лише частково розщеплюється за допомогою бактерій у товстому відділі кишок до дисахариду целобіози і моносахариду глюкози. Тут же мікроорганізми використовують клітковину для біосинтезу вітамінів групи К. Клітковина необхідна як стимулятор секреторної і моторної функцій травного каналу і для формування калу, тому вона є важливою складовою частиною харчового раціону людини. Проте надмірне надходження клітковини в організм викликає посилення перистальтики кишок, і їжа проходить через травний канал дуже швидко. У результаті цього харчові продукти не повністю перетравлюються і всмоктуються, значна частина їх потрапляє в кал і видаляється з організму невикористаною. Поряд із цим при підвищеній кількості клітковини в харчовому раціоні в товстому відділі кишок посилюються процеси бродіння, відбувається скупчення газів, що може призвести до серйозних ускладнень у хворих людей, а також у тих, хто мало рухається. Такі полісахариди, як крохмаль і глікоген, що надходять із їжею, попередньо підлягають розщепленню в травному каналі під дією певних ферментів. У результаті розщеплення утворюються моносахариди, які потім легко асимілюються тканинами організму. Процес перетравлювання вуглеводів (крохмалю і дисахаридів) починається вже в ротовій порожнині під дією двох ферментів, що містяться в складі слини, - амілази і а-глюкозидази (мальтази). Амілаза розщеплює крохмаль, а-глюкозидаза - мальтозу. Амілаза слини є більш активною порівняно з мальтазою, тому в ротовій порожнині з крохмалю утворюються переважно великі фрагменти його молекул - декстрини, що мають солодкий смак, і лише незначна кількість мальтози і глюкози.
Оскільки в ротовій порожнині їжа довго не затримується, подальше розщеплення вуглеводів продовжується в наступних відділах травного каналу. У шлунку немає ферментів, які розщеплюють вуглеводи. До того ж ферменти слини припиняють свою дію в шлунку, де реакція середовища кисла, а оптимум дії цих ферментів лежить у нейтральному або слабокислому середовищі. У дванадцятипалій кишці й у порожнині тонкої кишки під дією ряду ферментів підшлункової залози і кишкового соку відбувається остаточне розщеплення вуглеводів до моносахаридів. Так, а-амілаза розщеплює крохмаль і декстрини до мальтози, α-глюкозидаза розщеплює мальтозу на дві молекули глюкози. Під дією β-галактозидази (лактази) кишкового соку лактоза розщеплюється на глюкозу і галактозу, β-фруктофуранозидаза (сахараза) розщеплює сахарозу на глюкозу і фруктозу. Моносахариди, що утворилися, глюкоза, фруктоза, галактоза добре розчинні у воді - є тими простими цукрами, що всмоктуються стінкою кишок і через капіляри кишкових ворсинок потрапляють у кров'яне русло. Під час усмоктування частина моносахаридів (фруктоза, галактоза і маноза) перетворюються в глюкозу. У такий спосіб із кишок у кров потрапляє переважно глюкоза, і лише в незначних кількостях - фруктоза, галактоза і маноза. З током крові через ворітну вену вони потрапляють у печінку й інші органи і тканини. Частина глюкози затримується в печінці і використовується переважно для біосинтезу глікогену й інших складних вуглеводів, частина перетворюється в жири й окиснюється в тканинах. За нормальних умов на синтез глікогену використовується 3-5% глюкози, 30-35% її перетворюється в жири, а основна маса (60-70%) окиснюється до СО2 і H2О з виділенням енергії. Невеличка кількість глюкози є обов'язковою складовою частиною крові. За нормою це складає до 120 мг%. Цей рівень підтримується, незважаючи на постійне споживання глюкози тканинами і періодичне надходження її" з кишок після прийому їжі. Головна роль у підтримці постійної концентрації глюкози в крові належить печінці. За недостатньої кількості глюкози в крові під впливом нервових імпульсів, що йдуть із центральної нервової системи, частина глікогену печінки розщеплюється, підвищуючи тим самим рівень глюкози в крові. Цей процес називають мобілізацією глікогену. Регуляторну роль у підтримці постійного рівня глюкози в крові центральна нервова система здійснює не тільки шляхом прямого впливу на печінку, але й через ендокринні залози, серед яких найбільше значення мають надниркові і підшлункова залози. У мозковій речовині надниркових залоз утворюється адреналін, підвищений вміст якого в крові стимулює розщеплення глікогену в печінці. Підшлункова залоза виробляє гормон інсулін. Підвищене виділення цього гормону в кров супроводжується зниженням глюкози в крові. Таким чином, інсулін діє протилежно адреналіну: він затримує розпад глікогену в печінці і сприяє відкладенню його в м'язах.
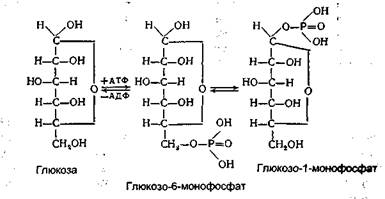
Крім інсуліну, в підшлунковій залозі виробляється ще один гормон - глюкагон, дія якого протилежна інсуліну. Істотний вплив на рівень глюкози в крові має і ряд інших гормонів: соматотропін, кортикостерон, тироксин. Всі ці гормони викликають підвищення вмісту глюкози в крові, через що їх називають діабетогенними (від назви захворювання "діабет", характерним для якого є підвищений вміст цукру в крові в результаті порушення функції підшлункової залози, що виробляє інсулін). Концентрація глюкози в крові може збільшуватися до 160-180 мг% при вживанні з їжею великої кількості вуглеводів. Це явище зветься аліментарною, або харчовою, гіперглікемією, що супроводжується виділенням глюкози із сечею, тобто глюкозурією. З вищевикладеного випливає, що надходження глюкози в кров відбувається переважно двома шляхами: розщепленням глікогену в печінці й усмоктуванням простих цукрів із кишок після прийому їжі. Утворення глюкози з глікогену відбувається не прямим його розщепленням, а за допомогою ферментів шляхом приєднання до нього фосфатної кислоти. При цьому від глікогену послідовно відщеплюються залишки глюкози у вигляді фосфорних ефірів, які потім розщеплюються на глюкозу і фосфатну кислоту:
Невелика кількість глюкози утворюється також шляхом гідролітичного розщеплення глікогену, проте цей шлях має дргорядне значення. БІОСИНТЕЗ ВУГЛЕВОДІВ Існує два основні шляхи біосинтезу вуглеводів із відносно нескладних метаболітів. Один із них полягає у відновленні вуглекислого газу до глюкози. Цей процес, характерний для зелених рослин і названий фотосинтезом, здійснюється за рахунок енергії сонячних променів за допомогою хлорофілу згідно з рівнянням:
Уловлюючи сонячні промені і перетворюючи їх енергію в енергію вуглеводів, зелені рослини забезпечують зберігання і розвиток життя на Землі. У цьому полягає, за словами К.А. Тимірязєва, космічна роль зелених рослин як посередника між сонцем і всім живим на Землі. Останнім часом роботи групи вчених Інституту біохімії ім. О.В. Палладіна НАН України під керівництвом академіка М.Ф. Гулого показали, що тканини вищих тварин також здатні фіксувати вуглекислий газ, хоча механізм його фіксації відрізняється від такого ж у фотосинтезуючих клітинах. Він полягає в нарощуванні вуглецевого скелета оксидом вуглецю (IV) таких субстратів, як кетокислоти, жирні кислоти, амінокислоти й ін. У печінці, нирках і скелетних м'язах людини І вищих тварин існує інший шлях біосинтезу вуглеводів, який називають глюконеогенезом. Це синтез глюкози з піровиноградної або молочної кислоти, а також із так званих глікогенних амінокислот, жирів і інших попередників, що в процесі метаболізму можуть перетворюватися в піровиноградну кислоту або метаболіти циклу трикарбоновых кислот. Глюконеогенез - це шлях, обернений гліколізу. Проте цей шлях має три стадії, що в енергетичному відношенні не можуть бути використані при перетворенні піровиноградної кислоти в глюкозу. Ці три стадії гліколізу замінені "обхідними" реакціями з меншою витратою енергії. Першою обхідною реакцією є перетворення піровиноградної кислоти у фосфоенолпіровиноградну. Оскільки розщеплення глюкози відбувається в мітохондріях, а синтез - у цитоплазмі, на першому етапі мітохондріальна піровиноградна кислота перетворюється спочатку в щавлево-оцтову. Каталізує це перетворення фермент піруваткарбоксилаза, яка активується ацетил-КоА за участю АТФ. Щавлево-оцтова кислота, що утворилася, відновлюється потім за участю НАД Н + Н+ в яблучну:
Яблучна кислота дифундує в цитоплазму, окиснюється цитоплазматичною малатдегідрогеназою з утворенням цитоплазматичної щавлево-оцтової кислоти, з якої утворюється фосфоенол-піровиноградна кислота. Цю реакцію каталізує фосфоенолпіру-ваткарбоксикіназа. Донором фосфатної кислоти виступає ГТФ;
Далі відбувається ціла серія обернених реакцій, які закінчуються утворенням фруктозо-1,6-дифосфату. Перетворення фруктозо-1,6-дифосфату у фруктозо-6-фосфат - друга незворотна реакція гліколізу. Тому вона каталізується не фосфофруктокіназою, а фруктозодифосфатазою. Цей фермент каталізує незворотний гідроліз 1-фосфатної групи: Фруктозо-1,6-дифосфат + Н2О → Фруктозо-6-фосфат + Н3РО4 На наступній (оборотній) стадії біосинтезу глюкози фруктозо-6-фосфат перетворюється в глюкозо-6-фосфат під дією фосфоглюкоізомерази гліколізу. Розщеплення глюкозо-6-фосфату до глюкози - третя незворотна реакція, яка не здійснюється шляхом обернення гексокіназою. Вільна глюкоза утворюється за допомогою глюкозо-6-фосфатази, що каталізує реакцію гідролізу: Глюкозо-6-фосфат + Н2O → Глюкоза + Н3РO4. У більшості клітин глюкозо-6-фосфат, який утворюється в процесі глікогенолізу, використовується як попередник для біосинтезу оліго- і полісахаридів. Велику роль у біосинтезі цих складних сахаридів відіграє сполука уридиндифосфоглюкоза, яка виконує роль проміжного переносника глюкози. Під час біосинтезу глікогену, наприклад, глюкозо-6-фосфат, перетворившись у глюкозо-1-фосфат під дією фосфоглюкомута-зи, взаємодіє з уридинтрифосфатною кислотою (УТФ) - сполукою, аналогічною АТФ, в яку замість аденіну входить азотиста основа урацил. У результаті цієї взаємодії за допомогою глюкозо-1-фосфатуридилтрансферази утворюється уридилдифосфо-глюкоза:
На заключному етапі біосинтезу глікогену в реакції, що каталізується глікогенсинтетазою, залишок глюкози з УДФ-глюкози переноситься на кінцевий залишок глюкози амілазного ланцюга з утворенням 1,4-глікозидного зв'язку. Розгалуження глікогену шляхом утворення 1,6-зв'язків завершується аміло-1,4-1,6-трансглюкозидазою. Біосинтез глікогену здійснюється не тільки з глюкозо-6-фосфату, який утворився шляхом глюконеогенезу. Як уже зазначалося вище, для його біосинтезу використовується також частина глюкози після всмоктування. Синтез глікогену як процес утворення рухливого резерву вуглеводів в організмі має велике біологічне значення. Головна роль у цьому належить печінці. Завдяки синтезу і відкладанню глікогену в печінці підтримується постійна концентрація глюкози в крові й інших тканинах, а також запобігаються втрати її з сечею при вживанні їжі, особливо вуглеводної. Крім того, відкладання глікогену в печінці сприяє поступовому використанню вуглеводів залежно від умов існування організму. Використанню глюкози для синтезу глікогену передує утворення глкжозофосфорних ефірів. Спочатку утворюється глюкозо-6-монофосфат. Джерелом енергії і донатором фосфату є АТФ. Каталізує цю реакцію гексокіназа. Під дією ферменту фосфо-глюкомутази глюкозо-6-монофосфат перетворюється в глюкозо-1-монофосфат:
Подальше перетворення глюкозо-1-монофосфату в глікоген відбувається вже знайомим нам шляхом.
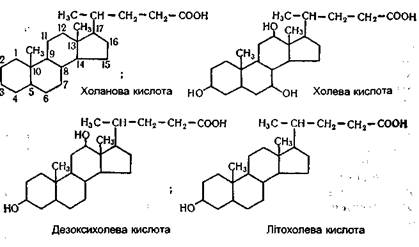
Обмін ліпідів Ліпіди являють собою велику групу органічних сполук. Всі вони різняться за своїм хімічним складом і структурою, але мають одну загальну для них властивість - нерозчинність у воді. Оскільки ферменти, які діють на ці органічні сполуки, водорозчинні, розщеплення й усмоктування ліпідів у харчовому каналі характеризуються деякими особливостями. Наявність же ліпідів різної структури зумовлює різноманітні шляхи їх розщеплення і синтезу. Зупинимося на обміні жирів, фосфатидів і стеридів, які мають найбільш важливе біологічне значення. Обмін ліпідів, як і вуглеводів, - багатоступінчатий процес, що складається з травлення, усмоктування, транспортування кров'ю, внутрішньоклітинного окиснення і біосинтезу. ТРАВЛЕННЯ ЛІПІ ДІВ Перетравлення тригліцеридів. Тригліцериди, або нейтральні жири, є концентрованими джерелами енергії в організмі. При окисненні 1 г жиру звільняється близько 38,9 кДж енергії. Як гідрофобні сполуки жири резервуються в компактній формі, займаючи порівняно мало місця в організмі. Разом із їжею в організм людини щодоби надходить до 70 г жирів рослинного і тваринного походження. За своєю хімічною природою вони є головним чином триглі церидами. Розщеплення жирів відбувається за допомогою ферментів, які називають ліпазами. Слина не містить таких ферментів, тому в ротовій порожнині жири ніяких змін не зазнають. У шлунку активність ліпази дуже слабка. Це викликано тим, що в шлунку реакція середовища дуже кисла (рН=1,5 -2,5), у той час як оптимум дії ліпази знаходиться при рН=7,8 - 8,1. У зв'язку з цим у шлунку перетравлюється всього 3-5% жирів, що надходять. Перетравлювання жирів у шлунку відбувається тільки в новонароджених і дітей грудного віку. Це пов'язано з тим, що рН середовища в шлунку новонароджених становить 5,6, а за цих умов ліпаза проявляє велику активність. Крім того, жир молока матері, який є основним продуктом харчування дітей у цей період, знаходиться в сильно емульгованому стані, а саме молоко містить ліполітичний чинник, що бере участь у перетравлюванні жирів. Проте шлунок усе не відіграє певну роль у процесі перетравлювання жирів у дорослих. Він регулює надходження жиру в кишки і перетравлює білки, звільняючи таким шляхом жир з ліпопротеїдних комплексів їжі. Основним місцем перетравлювання жирів є дванадцятипала кишка і відділи тонкої кишки. Оскільки жири нерозчинні у воді, а ферменти, що розщеплюють їх, є водорозчинними сполуками, необхідною умовою для гідролітичного розщеплення жирів на складові частини є їх диспергування (подрібнення) з утворенням тонкої емульсії. Диспергування й емульгування жиру відбувається в результаті дії декількох чинників: жовчних кислот, вільних вищих жирних кислот, моно- і дигліцеридів, а також білків. Цьому сприяють також перистальтика кишок і вуглекислий газ, що постійно утворюється при взаємодії кислих компонентів їжі, що надходять із шлунка, з карбонатами кишок, які створюють лужне середовище. Вуглекислий газ, що утворюється, "пробулькує" через харчові маси, беручи участь таким чином у диспергуванні жиру. Нейтралізації вмісту шлунку сприяє також надходження в просвіт тонкої кишки жовчі, яка має лужний характер. Жовч - густа рідина світло-жовтого кольору зі специфічним запахом, гірка на смак. До складу жовчі входять жовчні кислоти, жовчні пігменти, продукти розпаду гемоглобіну, холестерин, лецитин, жири, деякі ферменти, гормони й ін. Жовч сприяє перистальтиці тонкої кишки, здійснює бактеріостатичну дію на її мікрофлору. З жовчю виділяються з організму отрути. Вона є також активатором ліполітичних ферментів і підвищує проникність стінки кишок. Головною складовою частиною жовчі є жовчні кислоти. Вони утворюються в печінці з холестерину і знаходяться в жовчі як у вільному, так і зв'язаному стані, а також у вигляді натрієвих солей. У жовчі людини містяться в основному три жовчні кислоти. Основну масу складають холева (3,7,12-тригідрокси-холанова) і дезоксихолева (3,12-дигідроксихоланова), невелику частину - літохолева (3-гідроксихоланова) кислоти, які є похідними холанової кислоти:
Холева кислота може знаходитися в жовчі також у зв'язаному стані у вигляді парних сполук з гліцином і похідним цистеїну таурином- відповідно глікохолевої і таурохолевої кислот:
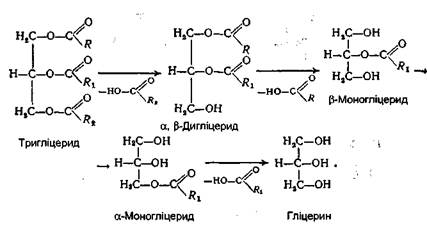
Завдяки наявності жовчних кислот відбувається зменшення поверхневого натягу ліпідних крапель, що сприяє утворенню дуже тонкої і стійкої емульсії, діаметр часток якої становить близько 0,5 мкм. Утворенню емульсії сприяють також моногліцериди і вищі жирні кислоти. Емульгування жиру призводить до колосального збільшення поверхні дотикання ліпази з водним розчином. Таким чином, чим тонша емульсія жирів, тим краще і швидше вони розщеплюються ліпазою. Крім того, у вигляді тонкої емульсії жири можуть навіть усмоктуватися стінкою кишок безпосередньо, не розщеплюючись на складові частини. За наявності жовчних кислот під дією ліпази в просвіті тонкої кишки відбувається гідролітичне розщеплення жирів. У результаті цього утворюються продукти часткового і повного розщеплення жирів - моно- і дигліцериди, вільні вищі жирні кислоти і гліцерин:
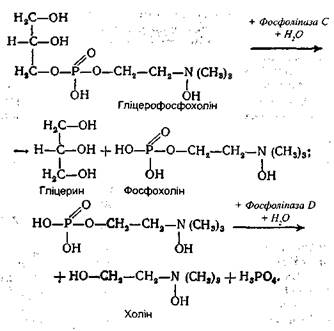
Тут же міститься і частина нерозщепленого жиру у вигляді дуже тонкої емульсії. Всі ці продукти надалі всмоктуються стінками кишок. У суміші тригліцериди складають близько 10%, моно- і дигліцериди - також 10%, а основна маса (близько 80%) продукти повного розщеплення жирів (гліцерин і вищі жирні кислоти). Перетравлення фосфогліцеридів. Основним місцем перетравлювання фосфатидів також є дванадцятипала кишка. Емульгування цих ліпідів відбувається під впливом тих же речовин, що і три гліцеридів. Проте гідролітичне розщеплення фосфатидів здійснюється під дією фосфоліпаз А, В, С і D. Кожний фермент діє на певний складноефірний зв'язок фосфоліпіду. Гідролітичне розщеплення, наприклад, лецитину відбувається в такий спосіб:
Такому повному розщепленню підлягає незначна частина фосфатидів, оскільки проміжні продукти розщеплення добре розчинні у воді і легко всмоктуються стінками кишок. До того ж фосфогліцериди легко утворюють емульсії, які також можуть усмоктуватися кишковою стінкою.
Перетравлення стеридів. Стериди, що входять до складу їжі, емульгуються під впливом тих же чинників, що і жири, після чого піддаються гідролітичному розщепленню до вільних стеринів і вищих жирних кислот. Цей процес здійснюється під дією ферменту холестеринестерази. УСМОКТУВАННЯ ЛІПІДІВ У результаті травлення жирів, фосфатидів, стеридів у просвіті тонкої кишки утворюється значна кількість продуктів їх часткового і повного гідролітичного розщеплення; моно- і дигліцериди, вищі жирні кислоти, стерини, азотисті основи, фосфатна кислота. Міститься також невелика кількість три-гліцеридів, які знаходяться в тонкоемульгованому стані. Всі ці продукти всмоктуються стінкою тонкої кишки. Такі продукти розщеплення, як жирні кислоти і холестерин, погано розчиняючись у воді, утворюють з жовчними кислотами водорозчинні комплекси - так звані холеїнові кислоти. Ці кислоти легко проникають в епітеліальні клітини стінки кишок, де розщеплюються на складові частини. Звільнені жовчні кислоти повертаються в просвіт кишок і знову використовуються для транспортування нерозчинних у воді продуктів розщеплення жирів. Частина продуктів розщеплення (гліцерин, гліцеринфосфатна кислота, азотисті основи) добре розчинні у воді і легко проникають в епітеліальні клітини. Фосфатна кислота всмоктується в клітини епітелію стінки тонкої кишки у вигляді натрієвих і калієвих солей. В основі всмоктування ліпідів лежить ряд складних фізико-хімічних і біологічних процесів, для здійснення яких витрачається енергія макроергічних зв'язків АТФ. В епітеліальних клітинах слизистої оболонки кишок із продуктів гідролітичного розщеплення, що всмокталися, знову синтезуються ліпіди. Проте цей ресинтез призводить до утворення специфічних жирів, характерних для даного організму. Для утворення нейтральних жирів використовуються вищі жирні кислоти, гліцерин, моно- і дигліцериди. Одночасно відбувається і синтез фосфатидів, для яких використовуються головним чином гліцеринфосфатна кислота, гліцериди і дигліцериди, а також у невеликій кількості моногліцериди. З холестерину і вищих жирних кислот утворюються стериди. В епітеліальних клітинах стінок кишок із синтезованих ліпідів, а також крапель тригліцеридів, що всмокталися, вітамінів (A, D, Е, К) і білків утворюються комплекси розміром 150-200 нм, які називаються хіломікронами, Хіломікрони наповнені різного роду ліпідами, утвореними переважно тригліцеридами, зовнішня білкова оболонка дозволяє хіломікронам добре розчинятися у воді. Хіломікрони дифундують спочатку в міжклітинну рідину, потім у лімфатичні капіляри і зрештою потрапляють у кров'яне русло, де під дією гепарину розпадаються на дрібні частинки. З током крові вони розносяться по всьому організму і відкладаються в резерв у жирових депо - під шкірою, навколо нирок, сальнику, брижі, м'язовій тканині. Частина жирів крові використовується для пластичних цілей, як джерело хімічної енергії і т.д. Таким чином, хіломікрони є переносниками утворених епітеліальними клітинами тонкої кишки ліпідів. При цьому вони транспортують у крові головним чином тригліцериди. Поряд з хіломікронами існують і інші форми транспорту ліпідів кров'ю, наприклад α- і β-ліпопротеїди. Найбільш рухливою формою ліпідів є вільні вищі жирні кислоти. Важлива роль в активному транспортуванні ліпідів належить форменим елементам крові. Еритроцити, наприклад, беруть участь у перенесенні фосфатидів і холестерину, лейкоцити - тригліцеридів. Велика роль в обміні ліпідів належить жировим депо. Дослідження показали, що в жирових депо відкладається не тільки знов синтезований в організмі специфічно видовий жир, але й у невеликих кількостях чужорідний, тобто той, що входив до складу їжі. Досліди, проведені на собаках, які голодували, показали, що харчові жири після всмоктування надходять спочатку в жирові депо, з яких переходять у плазму крові. Таким чином, жирова тканина не є пасивним депо жирів, склад її постійно поновлюється за рахунок ліпідів, що всмоктуються з кишок або синтезуються в організмі. БІОСИНТЕЗ ТРИГЛІЦЕРИДІВ Синтез нейтральних жирів складається з трьох процесів: утворення вищої жирної кислоти, утворення гліцерину І сполучення цих речовин у молекулу триглідериду. Синтез вищих жирних кислот. Місцем утворення вищих жирних кислот є цитоплазма. У процесі беруть участь активна форма оцтової кислоти, тобто ацетил-КоА, і вуглекислий газ. На першому етапі біосинтезу при взаємодії ацетил-КоА і СО2 утворюється проміжна сполука - малоніл-КоА. Утворення цієї сполуки відбувається за участі вітаміну Н з використанням енергії АТФ:
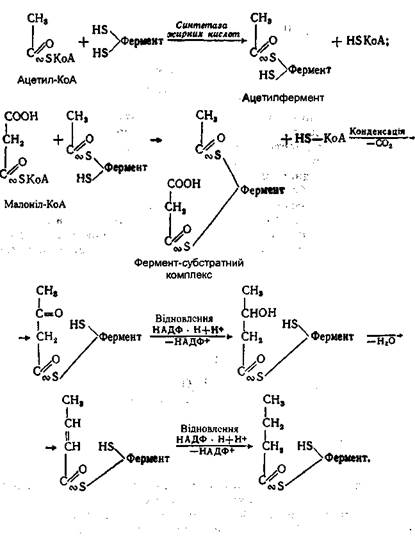
У процесі біосинтезу вищої жирної кислоти малоніл-КоА розщеплюється на ацетил-КоА і СО2, а окремі молекули ацетил-КоА з'єднуються між собою в довгий ланцюг. Отже, малоніл-КоА не входить до складу ланцюга вищої жирної кислоти, а служить лише проміжною формою, яка забезпечує утворення вищої жирної кислоти з окремих молекул ацетил-КоА. Весь процес з'єднання молекул ацетил-КоА здійснюється за допомогою ферменту синтетази жирних кислот, яка містить дві сульфгідрильні групи: центральну і периферичну. На центральній тіоловій групі здійснюється реакція конденсації між малоніл-КоА й ацетил-КоА з виділенням СО2 і відновлення утвореного продукту, а периферична група служить для утримання утвореного ланцюга:
У результаті з'єднання двох молекул ацетил-КоА і двох відновних реакцій утворюється фрагмент вищої жирної кислоти, який складається з чотирьох атомів вуглецю. На наступному етапі утворений фрагмент переноситься з центральної тіолової групи ферменту на периферичну:
При цьому центральна тіолова група звільняється і знову вступає в реакцію з новою молекулою малоніл-КоА. Потім у тій же послідовності відбувається реакція конденсації між утвореним ланцюгом і молекулою малоніл-КоА з виділенням С02 І реакції відновлення продукту, що утворився знову. Таким чином, при багаторазовому повторенні цих реакцій вуглеводний ланцюг усе більше подовжується, доки не з'єднає 16-18 атомів вуглецю. Після цього синтезована вища жирна кислота взаємодіє з молекулою коензима А, утворюючи активну форму у вигляді ацил-КоА й звільняючи при цьому синтетазу жирної кислоти:
Біосинтез тригліцеридів. Утворення жиру відбувається в результаті взаємодії молекули гліцерину з трьома молекулами вищих жирних кислот. Основним джерелом гліцерину в організмі є проміжний продукт окиснення вуглеводів - діоксіацетон-монофосфат, який шляхом відновлення перетворюється спочатку в гліцеринфосфатну кислоту, а потім у вільний гліцерин. Субстратом у біосинтезі жирних кислот, як ми тільки-но побачили, служить активна форма оцтової кислоти - ацетил-КоА. Як показали дослідження, гліцерин вступає б реакцію у вигляді гліцеринфосфату, а жирні кислоти - у вигляді своєї активної форми - ацил-КоА. На першому етапі біосинтезу триглі-церидів відбувається утворення фосфатидної кислоти - загального проміжного продукту в біосинтезі жирів І фосфатидів: і Далі фосфатидна кислота ферментативним шляхом розщеплюється на фосфатну кислоту і α, β-дигліцерид, що також є загальним проміжним продуктом у біосинтезі жирів і фосфатидів:
Утворений α, β-дигліцерид взаємодіє з третьою молекулою ацилкоензиму А, утворюючи молекулу тригліцериду:
Синтезований таким шляхом специфічний для організму жир відкладається в жирових депо. Біосинтез лецитину. Лецитин, як відомо, належить до групи фосфатидів. Для його біосинтезу необхідні такі речовини: гліцерин, вищі жирні кислоти, фосфатна кислота й азотиста основа холін. За своєю будовою лецитин деякою мірою нагадує триглі-цериди. Тому багато етапів у біосинтезі лецитину аналогічні етапам біосинтезу нейтральних жирів. Як уже було сказано, загальною проміжною сполукою в біосинтезі фосфатидів і тригліцеридів є α, β-дигліцерид. Саме до цієї проміжної сполуки і приєднуються фосфатна кислота і холін. Відбувається це в такий спосіб. Спочатку холін фосфорилюється за допомогою АТФ з утворенням фосфохоліну:
який далі взаємодіє з цитидинтрифосфатною кислотою (ЦТФ) і перетворюється в цитидиндифосфохолін:
При взаємодії цитидинфосфохоліну з α, β-дигліцерндом утворюється лецитин:
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 450; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.148.102.90 (0.095 с.) |