Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
По нервному волокну, в синапсах и нервных центрахСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Возникшее возбуждение передается по структурам рефлекторной дуги и достигает эффекторного органа. В передаче возбуждения можно выделить два процесса: передача по нервному волокну – до его окончания и синаптическая передача – с окончания нервного волокна на другую клетку. Проведение импульса через синапс существенно отличается от проведения по нервному волокну тем, что в процесс вовлекается химическое вещество и рецепторы к нему. С л о в а р ь т е м ы: Синапс Пресинаптическая и постсинаптическая мембрана Медиаторы Мембранные рецепторы Синаптоактивные вещества Парасимпато- и симпатолитики и миметики Агонисты и антагонисты медиаторов Проведение возбуждения по нервному волокну Механизм проведения возбуждения в нервных волокнах объясняется возникновением локальных токов между возбужденными и невозбужденными участками нервного волокна. При возбуждении активируются натриевые каналы, и возникает ток действия (ПД). Мембрана аксона на всем протяжении омывается солевым раствором, который хорошо проводит ток. Поскольку рядом с деполяризованным участком мембраны расположен участок с нормальным распределением зарядов (поляризованный), возникает разность потенциалов, способная деполяризовать мембрану до КУД и на соседнем участке возникает ПД (рис. 13). Электрический ток действует как раздражитель, деполяризующий невозбужденный участок до КУД и приводящий к возникновению ПД в соседнем участке. В безмиелиновых волокнах возбуждение распространяется непрерывно, а в миелинизированных- «скачками» между перехватами Ранвье (сальтаторный тип проведения). Дело в том, что миелин оболочки делает ее не проницаемой для электрического тока, поэтому ее сопротивление почти в 1000 раз больше, чем в перехватах. Следовательно, деполяризация до КУД и развитие ПД может возникнуть только в местах перехватов – нервный импульс делает «скачок» сразу на большое расстояние и скорость его распространения существенно увеличивается (рис 14). Такое проведение возбуждения называется сальтаторное.
Рисунок 13 Проведение возбуждения по нервному волокну
Рисунок 14 Сальтаторное проведение импульса по миелинизированному волокну Закономерности проведения возбуждения по нервному волокну: 1. Возбуждение может распространяться в любом направлении, потому что соседние участки с обеих сторон электроотрицательны по отношению к возбужденному участку. 2. Возбуждение распространяется не затухая, т.к. разность потенциалов деполяризует участок мембраны до КУД, следовательно, возникает ПД стандартной величины. 3. Скорость проведения возбуждения тем больше, чем выше амплитуда потенциала действия, потому что больше возникающая на соседних участках аксона разность потенциалов. 4. Скорость проведения прямо пропорциональна диаметру нервного волокна. 5. Частота импульсов не изменяется. 6. Возбуждение проводится изолированно по каждому нервному волокну.
Проведение возбуждения в синапсах. Передача нервного импульса с нервного волокна на другую клетку осуществляется посредством СИНАПСОВ. Синапс - специализированная зона контакта между аксоном и другим нейроном, мышечной или секреторной клеткой, обеспечивающая передачу нервного импульса с нервного волокна на эффекторную клетку.
Рисунок 15 Зоны синаптических контактов отмечены стрелками Существуют синапсы с химическим и электрическим способами взаимодействия. Электрических синапсов в организме человека немного, плотные контакты между клетками обеспечивают такой же вариант передачи импульса, как и в нервных волокнах – с помощью возникающих в месте контакта местных токов. Мы рассмотрим только химические синапсы. В химических синапсах возбуждение передается с помощью химического вещества - МЕДИАТОРА.
Рисунок 16 Структура синапса
В структуре синапса принято выделять пресинаптическую мембрану, образованную терминалью аксона, синаптическую щель и постсинаптическую мембрану, каждая структура выполняет свою функцию. Пресинаптическая часть, терминаль аксона представлена, как правило, утолщением, в котором содержится множество гранул диаметром от 20- 40 до 120 нм. С помощью гистохимических методов было установлено, что в гранулах содержится химические вещества, различные для различных нейронов, эти химические вещества были названы медиаторами – химическими «посредниками» в передаче электрического импульса с пресинаптической к постсинаптической мембране. Медиаторы могут быть биогенными аминами: норадреналин, дофамин, гистамин, серотонин; аминокислотами: глутаминовая кислота, глицин; олигопептидами: вещество Р, энкефалины, эндорфины; метаболитами: аденозин, АТФ, АДФ; широко распространен и в центральных структурах, и на периферии медиатор ацетилхолин. ХИМИЧЕСКИЕ МЕДИАТОРЫ являются веществами, синтезируемыми в цитоплазме нейронов. Нейроны могут: 1) синтезировать один медиатор, 2) транспортировать по аксону с помощью микротрубочек, 3) упаковывать медиатор в гранулы, 4) хранить эти гранулы в терминалях аксонов, 5) освобождать медиатор при возбуждении, 6) захватить назад в терминаль аксона медиатор или его фрагменты, 7) восстановить гранулы и вновь заполнить их медиатором. Медиатор выделяется в синаптическую щель размером 10-50 мкм, и взаимодействует с мембранным рецептором постсинаптической мембраны. В синаптической щели возможно разрушение медиатора специфическими для каждого медиатора ферментами, метаболиты возвращаются в нейрон для ресинтеза. Такими ферментами могут быть ацетилхолинэстераза – фермент для разрушения ацетилхолина, моноаминооксидаза (МАО) – для разрушения норадреналина или дофамина. Возможен обратный захват не разрушенного медиатора терминалью аксона. Постсинаптическая мембрана имеет мембранные рецепторы – сложные комплексы интегральных белков, пронизывающих клеточную мембрану, как правило, состоящие из нескольких субъединиц. В мембранных рецепторах одна часть способна распознавать собственный медиатор и связываться с ним, а вторая – представлена или ионным каналом, или системой специальных мембранных ферментов, которые называются внутриклеточные посредники (рис.17).
Если взаимодействие медиатора с рецептором приводит к активации ионных каналов, а результатом является изменение потенциала мембраны, то такие рецепторы называются ионотропными. Если запускается система вторичных внутриклеточных посредников, происходит изменение активности ферментов и темпа метаболических процессов (гликолиз, липолиз), то такие рецепторы называются метаботропными (рис 18).
Рисунок 18 Ионотропные и метаботропные рецепторы
Рисунок 19 выделение медиатора и взаимодействие с рецепторами
Для выделения медиатора необходимо сочетание двух событий: 1) волна деполяризации (ПД), достигшая терминали аксона, эта волна обусловлена активацией быстрых натриевых каналов и 2) открытие медленных, потенциал-зависимых кальциевых каналов пресинаптической мембраны. Поступление кальция в клетку (по градиенту концентрации) стимулирует освобождение гранул с медиатором. Гранулы выделяются путем экзоцитоза, поэтому медиатор появляется в синаптической щели не отдельными молекулами, а квантами, примерно из нескольких тысяч молекул.
Рисунок 20 Этапы выделения медиатора I) Деполяризация пресинаптической мембраны и поступление ионов натрия в терминаль аксона II) Активация потенциалзависимых медленных кальциевых каналов и поступление ионов кальция в клетку III) Освобождение медиатора, диффузия его к постсинаптической мембране и взаимодействие с мембранными рецепторами IV) Судьба медиатора в синаптической щели 1) Возвращение метаболитов в терминаль аксона 2) Разрушение медиатора специфическими ферментами 3) Обратный захват медиатора
Результат взаимодействия медиатора с мембранными рецепторами постсинаптической мембраны приводит к изменениям проницаемости этой мембраны для ионов путем активации или инактивации ионных каналов. На постсинаптической мембране могут открываться (или закрываться) натриевые, кальциевые калиевые или хлорные каналы. Результатом этого процесса будет изменение мембранного потенциала постсинаптической мембраны. Если деполяризующий ток ионов велик (натриевые каналы), на постсинаптической мембране возникает импульс возбуждения – потенциал действия. Чаще же на постсинаптической мембране возникает лишь незначительная, не достигающая порогового уровня деполяризация, локальный ответ (ЛО). В зависимости от расположения синапса этот ЛО называется или ВПСП (возбуждающий постсинаптический потенциал в центральных синапсах) или ПКП (потенциал концевой пластинки в нервномышечных синапсах).
Рисунок 21 Изменение потенциалов пресинаптической и постсинаптической мембраны 1 – пресинаптическая мембрана, 2 – постсинаптическая мембрана
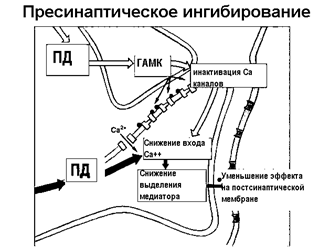
Синапсы можно классифицировать по локализации: на центральные и периферические. Центральные – это синапсы между нейронами, они отличаются местом взаимодействия терминали пресинаптического нейрона с постсинаптическим и могут быть аксо-соматическими, аксо-аксональными, аксо-дендритическими и дендро-дендритическими, кроме того, бывают аксо-вазальные синапсы, в которых медиатор выделяется в кровь. Периферические – это синапсы нервно-мышечные и нервно-секреторные, которые могут быть и соматическими и вегетативными, симпатические и парасимпатические. В периферических синапсах представлено два основных медиатора: ацетилхолин и норадреналин. Очень широкие возможности модулировать (видоизменить) работу синаптического контакта открывает наличие в ЦНС аксо-аксональных синапсов. Такие синапсы, в которых выделяются другие медиаторы, могут или «помочь» выделению медиатора по основному пути (рис. 22Б), или «помешать», ослабить выделение медиатора (рис.22 А).
Рисунок 22 Взаимодействие синапсов
Мембранные рецепторы тоже можно классифицировать и по веществу, с которым они взаимодействуют, и по эффекту такого взаимодействия. Рецепторы, взаимодействующие с ацетилхолином (АХ) называются холинорецепторами. В функциональном отношении они разделяются на две группы: М- и Н-холинорецепторы. М - чувствительные к мускарину, Н - чувствительные к никотину. В синапсах скелетных мышц присутствуют только Н-холинорецепторы, а в гладких мышцах внутренних органов - преимущественно М-холинорецепторы. Рецепторы, взаимодействующие с норадреналином называются адренорецепторами, и делятся на альфа- и бета- адренорецепторы. В постсинаптической мембране гладкомышечных клеток внутренних органов и кровеносных сосудов часто соседствуют оба вида адренорецепторов. Действие норадреналина является деполяризующим, если он взаимодействует с альфа-адренорецепторами и гиперполяризующим при взаимодействии с бета-адренорецепторами (таблица 6). Холин- и адрено - реактивные структуры находятся во всех внутренних органах, железах внутренней и внешней секреции, скелетной и гладкой мускулатуре, вегетативных ганглиях и ЦНС. Обратите внимание на то, что чувствительность адренорецепторов к адреналину и норадреналину различна, поэтому можно наблюдать несколько различные эффекты при активации симпатической нервной системы и при повышении в крови адреналина. Таблица 6
|
||||||
|
|
Последнее изменение этой страницы: 2016-08-14; просмотров: 973; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.59.36.4 (0.01 с.) |






 Рисунок 17 Кооперированный с натриевым каналом мембранный рецептор к самому распространенному медиатору ЦНС – глутамату. 1,2,3 – медиаторы, или похожие на них вещества, которые могут быть распознаны рецептором и связаны с ним, 4 – ион натрия, 5 – ворота ионного канала
Рисунок 17 Кооперированный с натриевым каналом мембранный рецептор к самому распространенному медиатору ЦНС – глутамату. 1,2,3 – медиаторы, или похожие на них вещества, которые могут быть распознаны рецептором и связаны с ним, 4 – ион натрия, 5 – ворота ионного канала