Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
По биологии индивидуального развитияСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
ПРАКТИКУМ ПО БИОЛОГИИ ИНДИВИДУАЛЬНОГО РАЗВИТИЯ Минск, 2010 ЛАБОРАТОРНОЕ ЗАНЯТИЕ № 1. СТРОЕНИЕ И ФУНКЦИЯ ПОЛОВЫХ КЛЕТОК. Цель работы: закрепить знания по строению гамет, этапам гаметогенеза и физиологии процессов, происходящих на этих этапах. Дать характеристику и объяснение процессам, происходящим в мейотическую фазу развития половых клеток.
Оборудование, приборы и принадлежности: таблицы, микроскопы, микропрепараты, схемы, планшеты.
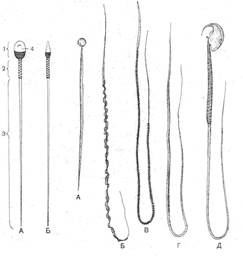
I. Теоретическая часть. Строение половых клеток. Половыми клетками (гаметами) являются мужская половая клетка сперматозоид и женская половая клетка – яйцеклетка. Половые клетки гаплоидны, т.к. образуются в результате редукционного деления мейоза. 1.1. Сперматозоид имеет все структуры, необходимые для обеспечения процесса оплодотворения: гаплоидное ядро с генетической информацией, заключенной в хромосомах, акросому – мембранную органеллу с ферментами, обеспечивающими проникновение ядра в яйцеклетку и двигательную систему, обеспечивающую перемещение ядра. Двигательный аппарат у различных биологических видов различен, что зависит от условий окружающей среды. Так, у некоторых видов (например, аскариды) сперматозоид перемещается при помощи амебоидного движения. У большинства видов передвижение осуществляется благодаря биению жгутиков, преобразующих химическую энергию АТФ в механическую за счет ферментов (рис. 1.1).
Рис. 1.1. Сперматозоиды позвоночных: I. А, Б – человека. II. А – щуки; Б – саламандры; В – голубя; Г – ехидны; Д – полевой мыши.
Жгутиковый сперматозоид состоит из головки и хвостика, в котором различают связующий, промежуточный, главный и дистальный отделы. На обычных гистологических препаратах ядра сперматозоидов обладают резко выраженной базофилий и имеют вытянутую грушевидную форму. Ядро характеризуется плотным расположением хроматина, в котором наряду с основными гистоновыми, обладающими положительным зарядом белками, находятся также негистоновые белки. ДНК хромосом в хроматине плотно упакована, поэтому ядро имеет кристаллоподобную структуру, его объем сведен до минимума. Ядерная оболочка сперматозоида полностью лишена ядерных пор. В передней части ядра под цитолеммой сперматозоида находится акросома - производное комплекса Гольджи и аналог лизосомы, которая содержит ферменты, расщепляющие компоненты оболочек яйцеклетки. Цитоплазма сперматозоида редуцирована до минимума и очень тонким слоем покрывает ядро. Связующий отдел хвостика содержит проксимальную центриоль, которая прилежит к ядру, располагаясь в углублении ядерной оболочки. Здесь же находится дистальная центриоль. От нее отходит осевая нить - аксонема, имеющая типичную структуру жгутиков и состоящая из 9 периферических дуплетов микротрубочек и двух расположенных в центре одиночных микротрубочек. Аксонема продолжается во все отделы хвостика, редуцируясь в дистальном отделе. Снаружи напротив каждого дуплета дистальной центриоли в связующем отделе находится одна так называемая сегментированная колонна (всего их число равно 9). В промежуточном отделе хвостика сегментированные колонны продолжаются в 9 плотных волокон. В этом же отделе вокруг аксонемы и плотных волокон в виде спирали располагаются митохондрии. В главной части хвостика два из девяти плотных волокон, расположенные напротив друг друга, резко утолщаются и превращаются в продольные столбы, которые соединяются между собой боковыми ребрами. Формируется внешняя волокнистая оболочка, придающая жесткость и упругость хвостику. Таким образом, сегментированные колонны, плотные волокна, продольные столбы и соединяющие их ребра, а также внешняя волокнистая оболочка образуют каркас хвостика. В дистальном отделе количество микротрубочек сильно редуцируется. Снаружи хвостик покрыт цитолеммой. 1.2. Яйцеклетка – женская половая клетка с гаплоидным набором хромосом. При электронномикроскопическом исследовании в цитоплазме яйцеклетки выделяются митохондрии, комплекс Гольджи, хорошо развитые гранулярную и агранулярную эндоплазматическую сеть. Снаружи яйцеклетка окружена плазматической мембраной - цитолеммой, под которой располагается толстый слой цитоплазмы (кортикальный слой), содержащий кортикальные гранулы (рис. 1.2). Кортикальной слой играет важную роль в организации яйцеклетки, а также в оплодотворении.
Рис. 1.2. Электронная микрофотография овариального яйца крысы; в кортикальном слое видны кортикальные гранулы (1), микроворсинки яйца (2) и отростки (3) фолликулярных клеток (4), проникающие через zona pellucida
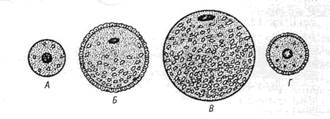
Яйцеклетки имеют хорошо развитый и своеобразно организованный цитоскелет. Его компоненты связаны с цитолеммой и вызывают постоянную модификацию поверхности клетки, в которой могут появляться и исчезать микроворсинки, меняться локализация рецепторов. Яйцеклетки окружены блестящей оболочкой и слоем фолликулярных клеток. Между блестящей зоной и цитолеммой овоцита имеется небольшое перивителлиновое пространство, которое существенно увеличивается после оплодотворения, поскольку в него выделяются содержимое кортикальных гранул. Фолликулярные клетки через отверстия в блестящей оболочке посылают к цитолемме овоцита свои отростки. В цитоплазме яйцеклетки содержатся запасные питательные вещества: жир, гликоген, желток. Желток представляет собой совокупность химических веществ (белков, жиров, углеводов) для снабжения развивающегося зародыша питательными веществами. Количество желтка в яйцеклетках сильно варьирует в разных группах животных. У амфибий, рептилий, птиц компоненты желтка синтезируются в печени, с током крови поступают в яичник, проходят через фолликулярный эпителий и поступают в овоцит путем микропиноцитоза. В ряду позвоночных в зависимости от наличия желтка, его количества и распределения по ооплазме различают алицетальные (без желтка) и лецитальные (с желтком) яйцеклетки (рис. 1.3).
Рис. 1.3. Основные типы яйцеклеток, различающиеся между собой по количеству и характеру распределения гранул желтка: А – олиголецитальная, первично изолецитальная яйцеклетка (ланцетник); Б – полилецитальная, умеренно телолецитальная яйцеклетка (амфибии); В – полилецитальная, резко телолицетальная яйцеклетка (птицы); Г – олиголецитальная, вторично изолецитальная яйцеклетка (млекопитающие). Лецитальные клетки делятся на олиголецитальные (маложелтковые) и полилецитальные (многожелтковые). В зависимости от распределения желтка по ооплазме яйцеклетки делятся на изолецитальные, умеренно телолецитальные и резко телолецитальные. В изолецитальных яйцеклетках желток распределен равномерно. В умеренно телолецитальных яйцеклетках желток находится на одном полюсе, который называется вегетативным. На другом полюсе - анимальном - лежат органеллы и ядро. В резко телолгцитальных клетках вегетативный полюс выражен особенно сильно и занимает большую часть клетки. Яйцеклетки млекопитающих, в том числе и человека, являются олигоизолецитальными: содержат очень мало желтка, который равномерно распределен по ооплазме. Кроме желтка цитоплазма яйцеклетки содержит морфогенетические факторы – молекулы, направляющие дифференцировку клеток для образования различных тканей и органов, которые рассеяны по всей яйцеклетке и распределяются между разными бластомерами при дроблении. 2. Прогенез (гаметогенез) – процесс формирования гамет. Гаметогенез делится на сперматогенез (образование сперматозоидов) и овогенез (образование яйцеклеток). 2.1. Сперматогенез – процесс формирования и развития мужских половых клеток - сперматозоидов. В сперматогенезе различают 4 фазы: размножение, рост, созревание и формирование. На протяжении периода размножения мужские половые клетки представлены сперматогониями. Это мелкие округлые клетки, делящиеся путем митоза. Они подразделяются на темные и светлые сперматогонии. Темные сперматогонии являются истинными стволовыми клетками, устойчивы к действию вредных факторов и редко делятся. Светлые сперматогонии разделяются на А и В-сперматогонии. А-сперматогонии являются полустволовыми клетками, способными к частым митотическим делениям. При делении каждой такой клетки могут возникать либо две А-сперматогонии, либо одна А- и одна В-сперматогония. В-сперматогонии также способны делиться митозом, но при этом не происходит полной цитотомии, и клетки оказываются связанными между собой цитоплазматическими мостиками. Возникают клоны (ассоциации) клеток. После некоторой паузы В-сперматогонии вступают в период роста, в течение которого превращаются в сперматоциты I порядка. Для периода роста характерно увеличение ядра и цитоплазмы развивающихся клеток, их размеры увеличиваются в четыре и более раз. Диплоидные сперматоциты I порядка не делятся митозом, но вступают в период созревания, который состоит из двух последовательных делений мейоза (мейоз I и мейоз II). Гаплоидные сперматоциты II порядка образуются в результате I деления мейоза. Мейоз I (редукционное деление) – в ходе его происходит уменьшение в два раза количества хромосом с формированием гаплоидного генома. Мейоз I имеет сложную профазу, состоящую из 5 стадий: лептотены, зиготены, пахитены, диилотены и диакинеза. На стадии лептотены хромосомы спирализуются и становятся видны в виде длинных тонких нитей. На стадии зиготены гомологичные хромосомы коньюгируют друг с другом. На стадии пахитены хромосомы сильно спирализуются и укорачиваются. В это время в них происходит кроссинговер - обмен генами. Диплотена характеризуется расщеплением хромосом на хроматиды и образованием тетрад. В диакинезе хромосомы еще более утолщаются и несколько отходят друг от друга. Таким образом, в профазе мейоза 1 происходит подготовка к редукции числа хромосом. В метафазе гомологичные пары хромосом располагаются на экваторе клетки. В анафазе к полюсам отходят целые гомологичные хромосомы, т.е. осуществляется редукция генома. В телофазе происходит цитотомия, в результате чего образуются два сперматоцита II порядка с гаплоидным набором хромосом. Их хромосомы состоят из двух хроматид. Мейоз II (эквационное или уравнительное деление) протекает по типу обычного митоза, но в гаплоидной клетке. В анафазе к полюсам отходят половинки хромосом–хроматиды, а в результате телофазы образуются сперматиды, содержащие хроматиды хромосом. Сперматиды, как и сперматоциты II порядка, содержат гаплоидный набор хромосом. Из I сперматоцита I порядка образуется 4 сперматиды. Все образующиеся в процессе сперматогенеза клетки (В-сперматогонии, сперматоциты I и II порядка, а также сперматиды) остаются связанными между собой цитоплазматическими мостиками в клеточные ассоциации или клоны. Окончательное разделение клеток происходит в фазу формирования. Сохранение цитоплазматических мостиков между клетками имеет большой биологический смысл. Оказывается, что для полноценной дифференцировки сперматозоидов необходим весь диплоидный геном и продукты его деятельности. Во-первых, потому, что в исходном диплоидном геноме могут содержаться дефектные, летальные аллели генов, и клетка, получившая их, погибнет, если не будет обеспечена продуктами нормального аллеля, находящегося в ядрах других клеток, его получивших. Во-вторых, как известно, одни мужские половые клетки получают Х-, другие - Y- половую хромосому. Каждая из них содержит много важных генов, необходимых для развития сперматозоидов. Поэтому, благодаря цитоплазматическим мостикам развивающиеся мужские половые клетки получают продукты деятельности всего диплоидного генома. Фаза формирования является самой продолжительной фазой сперматогенеза. В процессе ее из сперматид образуются сперматозоиды. Часто эту фазу называют спермиогенезом (рис. 1.4). Процесс спермиогенеза начинается образованием из комплекса Гольджи вначале акробласта, а затем акросомы, которая содержит ферменты для разрушения яйцевых оболочек. Центросома, состоящая из двух центриолей, перемещается на противоположный полюс. Проксимальная центриоль прилежит к ядру, а дистальная делится на две части. Из одной части формируется жгутик, который превращается в осевую нить хвостика. Вторая часть играет роль базального тельца. Образуются элементы цитоскелета: сегментированные колонны, плотные волокна, продольные столбы с ребрами.
Рис. 1.4. А—Ж — последовательные фазы спермиогенеза: 1 — ядро сперматиды; 2 — аппарат Гольджн; 3 — центриоли; 4 — зачаток акросомы; 5 — митохондрии; 6—жгутик; 7— акросома; 8 — проксимальная центриоль; 9 — дистальная центриоль; 10 — ядерное вещество, сосредоточенное в головке сперматозоида; 11-—митохондриальная спираль; 12 —остатки цитоплазмы; 13 — головка; 14 — шейка; 15 — средняя часть; 16 — хвост; 17 — концевой участок хвоста.
Цитоплазма сперматозоида сильно редуцируется, а ядро становится вытянутым, компактным и гипербазофильным. На заключительных этапах формирования сперматозоиды отделяются от соединяющей их друг с другом общей цитоплазмы и становятся свободными. Оставшаяся после отделения цитоплазма (остаточные тельца) подвергается фагоцитозу. 2.2. Овогенез протекает сходно со сперматогенезом, но имеет ряд отличий. Исходными клетками для овогенезя являются первичные половые клетки (гонобласты), развивающиеся в раннем эмбриональном периоде в женской половой железе – яичнике (рис. 1.5). Гонобласты превращаются в овогонии. Эти мелкие клетки вступают в фазу размножения и интенсивно делятся митозом. Затем все они превращаются в овоциты I порядка, которые блокируются на определенной стадии мейоза I. Далее овоциты I порядка вступают в длительный период роста. Период роста делится на две части: период малого или медленного роста, и период большого или быстрого роста. В период быстрого роста идет подготовка к прохождения мейоза. Третья фаза овогенеза - созревание - начинается перед овуляцией. Происходит первое мейотическое деление и образуется гаплоидный овоцит II порядка, который вступает во второе мейотическое деление, но блокируется в метафазе II.
Рис. 1.5. Схематическое изображение яичника: 1 — мезоварий; 2 —герминативный эпителий; 3 — первичный фолликул; 4 — двуслойный фолликул; 5 — начало образования полости фолликула; 6—атретический фолликул; 7— почти полностью созревший фолликул; 8 — атретический фолликул; 9 — зрелый фолликул; 10 — ооцит; 11 — полость, наполненная фолликулярной жидкостью; 12 — разорвавшийся фолликул; 13 — высвободившаяся яйцеклетка; 14 — развивающееся желтое тело; 15 — соединительная ткань; 16 — лютеиновые клетки; 17 — фибрин крвяного сгустка; 18 – свернувшаясякровь; 19 – соединительная ткань яичника; 20 – полностью сформировавшееся желтое тело; 21 – беловатое тело яичника; 22 – кровеносные сосуды.
Завершение созревания инициируется оплодотворением. В отличие от сперматогенеза, возникающие в ходе овогенеза в результате двух делений мейоза клетки, не равны по размерам. Из овоцита I порядка образуется крупный овоцит II порядка и очень мелкое направительное (редукционное) тельце. Первое редукционное тельце во II делении мейоза также может делиться на два направительных (редукционных) тельца. Следовательно, в результате двух делений мейоза образуется одна гаплоидная яйцеклетка и три гаплоидных направительных (редукционных) тельца, которые вскоре погибают. Таким образом, в процессе гаметогенеза формируются зрелые половые клетки. Генеративные клетки отличаются от соматических следующими параметрами: - гаплоидным набором хромосом, а значит, невозможностью обычного размножения и обычного метаболизма; - резко измененным ядерно-цитоплазматическим отношением (в обычных соматических клетках ЯЦО 1:6- 1:10; у сперматозоида ЯЦО составляет 1;0,2 - 1:0,5; а у яйцеклетки- 1:500) - нормальный метаболизм клетки может осуществляться только при теснейшем взаимодействии ядра и цитоплазмы: у половых клеток очень необычный обмен веществ, он находится в состоянии депрессии. - половые клетки - это высокоспециализированные клетки, обладающие многими специальными, выработавшимися в процессе эволюции приспособлениями для выполнения специфических функций: встреча клеток, их соединение, защита зиготы. - из-за высокой специфичности и необычного обмена веществ эти клетки нуждаются в особых условиях обитания, защитных и питательных структурах (оболочки яйцеклетки). - женские клетки содержат питательные вещества для развития зародыша, имеют громадный объем цитоплазмы и поэтому неподвижны; мужские клетки в десятки и сотни раз мельче, имеют запас энергии и жгутик для перемещения к женской клетке.
II. Методические указания к выполнению лабораторной работы. 1. Ознакомиться с происхождением первичных половых клеток в онтогенезе по методическому пособию. 2. Изучить ход и фазы мейоза (планшет № 1.1). Зарисовать общую схему мейоза. 3. Изучить процесс сперматогенеза. 3.1. Зарисовать первое деление созревания (редукционное) и второе (эквационное) деление в мейозе сперматогенеза. Обозначить периоды профазы 1 мейоза (планшет № 1.1). 3.2. Изучить последовательные стадии сперматогенеза. Зарисовать таблицу «Мужской гаметогенез» (планшет № 1.2). Зарисовать схему сперматогенеза, обозначить фазы развития гамет и названия клеток каждой из фаз. 3.3. Ознакомиться с процессами, происходящими при спермиогенезе (планшет № 1.3 «Участок стенки эпителя семенных канальцев крысы», «Дифференцировка сперматиды сперматозоида»). Найти сперматогонии типа А и В, сперматоциты в начале редукционного деления и сперматиды. Зарисовать изменения клеток в ходе спермиогенеза. Дать характеристику сперматиды. 3.4. Изучить строение сперматозоида (планшет № 1.4). Зарисовать схему строения мужской половой клетки, обозначить акросому и части хвостика (связующая, промежуточная, главная, концевая). 3.5. Изучить под микроскопом и зарисовать препарат № 1. Дать его описание. Препарат № 1. Спермии морской свинки (рис. 1.6). Зрелые спермии морской свинки представляют типичный образец подобного рода клеток. Длина их около 100 мк, отчетливо выражена головка, шейка с лежащей в ней центросомой, хвостовая нить. Начните изучение препарата с малого, позже переходите на большее увеличение. Зарисовку сделайте при большом увеличении. На спермии морских свинок очень похоже строение сперматозоидов человека.
Рис. 1.6. Спермии морской свинки.
4. Изучить процесс овогенеза. 4.1. Изучить последовательные стадии овогенеза. Зарисовать таблицу «Женский гаметогенез» (планшет № 1.2). Зарисовать схему овогенеза, обозначить фазы развития и названия клеток каждой из фаз. 4.2. Изучить строение женской половой клетки (планшет № 1.5). Зарисовать схему строения и обозначить оболочки клетки (блестящая, лучистый венец). 4.3. Изучить под микроскопом и зарисовать препарат № 2. Дать его описание. Препарат № 2. Яйцеклетки лягушки (рис. 1.7). Препарат представляет собой срез яичника лягушки, на котором видны яйцеклетки на разных стадиях «большого роста». Уже при малом увеличении заметны клетки разнообразных размеров и окраски. Мелкие клетки окрашиваются в синий цвет гематоксилином, более крупные окрашиваются более интенсивно в красный цвет эозином. Различие окраски цитоплазмы связано с изменением физико-химической структуры яйцеклетки вследствие накопления в ее цитоплазме желтка. Параллельно этому увеличиваются размеры клеток. В крупных ядрах видны ядрышки и хроматиновые глыбки – компоненты расплывшихся хромосом. Нарисуйте несколько клеток на разных стадиях, подчеркнув неодинаковость размеров и окраски.
Рис. 1.7. Яйцеклетки на разных стадиях «большого роста» в яичнике лягушки.
4.4. Изучить под микроскопом и зарисовать препарат № 3. Дать его описание. Препарат № 3. Яичник кошки (рис. 1.8). В яичнике млекопитающих все половые клетки являются овоцитами I порядка, т.е. проходят период роста. Однако отдельные яйцеклетки находятся на разных фазах этого периода. При малом увеличении отметим, что орган явственно разделяется на две части: наружную, более плотную и темноокрашенную – корковое вещество, и центральную, более светлую – мозговое вещество. То и другое состоит из переплетения мышечных и соединительнотканных волокон, среди которых выделяются перерезанные в разных направлениях кровеносные сосуды, содержащие красные кровяные клетки. В корковом слое, недалеко от поверхности, залегают своеобразные группы клеток. Они состоят из одной более крупной клетки со светлым ядром и широким поясом цитоплазмы, окруженной рядом более мелких, тесно к ней прилегающих клеток. Крупная, центрально расположенная клетка является овоцитом, а окружающие ее мелкие – питающими фолликулярными клетками. 4.5. Изучить под микроскопом и зарисовать препарат № 4. Дать его описание.
Рис. 1.8. Яичник. 1 – зачатковый эпителий; 2 – белочная оболочка; 3 – корковое вещество; 4 – первичные (примордиальные) фолликулы; 5 – растущие фолликулы; 6 –пузырчатый фолликул (графов пузырек); 7 – полость пузырчатого фолликула, заполненная жидкостью; 8 яйцеклетка (овонит первого порядка); 9 – пузырчатый фолликул, в котором яйценосный бугорок и овоцит не попали в срез; 10 – желтое тело; 11 – атретические тела; 12 – мозговое вещество; 13 – соединительная ткань и кровеносные сосуды.
Препарат № 4. Яйцеклетки млекопитающих (рис. 1.9). Яйцеклетка, окруженная рядом фолликулярных клеток, образует примордиальный, или первичный, фолликул. По мере роста овоцита количество питающих его клеток увеличивается, они располагаются в несколько рядов, и фолликул становится многослойным. Далее в нем возникают пространства, наполненные питательной жидкостью, расслаивающей толщу фолликулярных клеток, и массивный фолликул постепенно превращается в пузырек. В готовом виде последний является образованием до горошины величиной и различим невооруженным глазом. Фолликулярные клетки в нем оттесняются плазмой к самым стенкам, и только вокруг яйцеклетки сохраняется их больше, причем это образование в виде выроста – яйценосный бугорок – вдается в полость пузырька. Такой пузырек называется граафовым. Описанные фазы перестройки фолликула – однослойный, многослойный, графов пузырек – образуют фолликулярный цикл яичника. Зарисовать нужно при большом увеличении примордиальный и многослойный фолликул, а при малом – начальную стадию развития и вполне сформировавшийся граафов пузырек.
Рис. 1.9. Яйцеклетка кошки: 1 – ядро; 2 – цитолплазма; 3 – клетка фолликулярного эпителия
5. Разобрать классификацию яйцеклетки по количеству и распределению в ней желтка (по методическому пособию). Изучить типы яйцеклеток и способы питания яйцеклеток. 6. Сдать отчет преподавателю и защитить его. III. Содержание отчета. Отчет должен быть представлен на отдельных листах формата А4 или в альбоме. Отчет должен содержать: 1. Цель работы. 2. Краткое описание теории сперматогенеза, овогенеза, строения гамет. 3. Результаты исследований (микроскопическое изучение препаратов) и их анализ (с указанием использованных микроскопов, их увеличения, других приборов и материалов). 4. Результаты выполнения индивидуального задания (определение и описание «слепого» препарата). 5. Выводы. Отчет на листе формата А4 сдается в конце работы преподавателю.
IV. Контрольные вопросы. 1. Какие части различают в сперматозоиде? 2. Что такое акросома и какую роль она играет? 3. Каковы принципы классификации яйцеклеток? 4. Дайте характеристику фаз прогенеза. 5. В чем отличие мужского и женского гаметогенеза?
Рекомендуемая литература. 1. А.В.Белоусов. Биология индивидуального развития, 1983. 2. Т.М.Студеникина. Основы общей эмбриологии, 1999. 3. О.В.Волкова. Атлас. Гистология, цитология, эмбриология, 1996. 4. С.Л.Кузнецов. Атлас эмбриологии, 2002. ЛАБОРАТОРНОЕ ЗАНЯТИЕ № 2.
ЛАБОРАТОРНОЕ ЗАНЯТИЕ № 3.
ЛАБОРАТОРНОЕ ЗАНЯТИЕ № 4.
НЕЙРОГЕНЕЗ.
Цель работы: изучить процесс нейруляции и образования нервной трубки; изучить процесс образования нервного гребня и дифференцировку клеток нервного гребня; дать сравнительную характеристику нейруляции у млекопитающих и других позвоночных. Оборудование, приборы и принадлежности: таблицы, микроскопы, микропрепараты, схемы, планшеты.
I. Теоретическая часть ЛАБОРАТОРНАЯ РАБОТА № 5. ФОРМИРОВАНИЕ ОРГАНОВ.
Цель работы: изучить развитие производных зародышевых листков; уметь объяснить механизмы формообразования; иметь представление об индукционных взаимодействиях в ходе тканевой дифференцировки.
Оборудование, приборы и принадлежности: таблицы, микроскопы, микропрепараты, схемы, планшеты.
I. Теоретическая часть. После завершения гаструляционных движений зародыш приступает к построению отдельных систем своих органов — к органогенезу. Если формирование центральной нервной системы позвоночных связано, как мы видели, с морфологическими перестройками, охватывающими почти весь зародыш, то дальнейший органогенез представляет собой более местный процесс. Зародыш постепенно разбивается на относительно независимо развивающиеся системы, которые и превращаются в органы или части тела (рис. 5.1).
Рис. 5.1. Схема органогенеза и гистогенеза у зародыша высшего позвоночного. 1 – кожная эктодерма; 2 – ганглиозная пластинка; 3 – нервная трубка; 4 – хорда; 5 – дерматом; 6 – миотом; 7 – склеротом; 8 – нефротом; 9 – мезотелий; 10 – висцеральный листок спланхнотома; 11 – кишечная энтодерма; 12 – мезенхима; 13 – внезародышевая энтодерма; 14 – эндотелий аорты; 15 – желточная энтодерма; 16 – целом. 1. Развитие производных энтодермы. Сформировавшаяся кишечная трубка позвоночных может быть разделена на три отдела: переднюю, среднюю и заднюю кишки. Наиболее сложно дифференцирована передняя кишка. Относительно долгое время она представляет собой слепой вырост, так как ротовое отверстие прорывается на сравнительно поздних стадиях. До этого времени передняя кишка успевает дифференцироваться на глотку, зачаток желудка и зачаток печени. Последний возникает из печеночного выроста. Кроме того, за счет материала передней кишки впоследствии формируются большая часть двенадцатиперстной кишки и поджелудочная железа. На более поздних стадиях развития из вентральной стенки передней кишки образуются зачатки легких. Они появляются непосредственно сзади глотки на вентральной стороне пищеварительного канала в виде парных выпячиваний. Одна из наиболее характерных черт зародышей всех позвоночных — образование в стенке глотки жаберных карманов, часть которых превращается в сквозные жаберные щели. Наибольшее число жаберных щелей формируется у низших позвоночных. Например, у круглоротых их 14. У зародышей амфибий закладываются 5 пар жаберных карманов, из которых 4 задних открываются во внешнюю среду в виде жаберных щелей, а передний так и остается слепым выростом. При прорыве жаберных щелей покровная эктодерма соединяется с энтодермой передней кишки. У зародышей амниот закладываются 4 пары жаберных карманов, из которых 3 передних на короткое время превращаются в жаберные щели, но впоследствии снова зарастают. Первая пара карманов превращается в евстахиевы трубы, соединяющие полости среднего уха с ротовой полостью. Из других жаберных карманов развиваются железы внутренней секреции: 3-я пара карманов дает начало парному зачатку зобной железы (тимусу), 4-я — паращитовидным железам. Все эти железы образованы преимущественно энтодермальной выстилкой жаберных карманов. Из энтодермы дна глотки в виде непарного выпячивания между 1-й и 2-й жаберными дугами формируется щитовидная железа. В промежутках между жаберными щелями закладываются хрящевые жаберные дуги, образующие висцеральный скелет. Они строятся из переместившихся сюда клеток нервного гребня (эта закладка описана ниже). Чем выше организация позвоночного, тем больше сокращается число жаберных дуг, которые в ходе развития подвергаются все большим преобразованиям. У круглоротых и акуловых рыб закладываются 7 пар дуг, у других рыб — 5 пар. Хрящи 1-й пары превращаются у рыб в челюстные дуги, 2-й — в подъязычную дугу. У наземных позвоночных за счет хрящей первых двух пар возникают, кроме того, слуховые косточки. За счет остальных дуг у высших позвоночных формируются хрящи трахей. Кроме того, клетки дорсальных концов жаберных дуг участвуют в образовании головных нервных ганглиев. В морфологической дифференцировке легких, печени и поджелудочной железы много общего: она сводится к последовательному ветвлению первоначальных зачатков — выступов кишечного эпителия — на все более тонкие выросты, вклинивающиеся в окружающую их мезенхиму. 2. Развитие производных эктодермы. 2.1. Развитие кожи и ее придатков. Кожа позвоночных развивается из двух зародышевых листков — эктодермы и мезодермы. Эмбриональная эктодерма сначала превращается в двухслойный, а затем в многослойный эпителий — кожный эпидермис. Его внутренний, прилежащий к мезодерме слой (ростковый, или мальпигиев) в течение всей жизни организма сохраняет функции камбия: в нем происходят клеточные деления, и вновь образующиеся клетки перемещаются во внешние слои эпидермиса, где дифференцируются. У всех наземных позвоночных дифференцированные клетки внешних слоев эпидермиса (кератоциты) синтезируют роговое вещество — белок кератин. Дифференцировка клеток, синтезирующих кератин, сопровождается их инактивацией, они заполняются кератином, уплощаются, теряют ядро и формируют поверхностный омертвевающий слой кожи. Мезодермальный слой кожи (дерма) образуется соединительнотканными клетками, происходящими из кожных листков сомитов (дерматомов). За счет деятельности клеток дермы формируются коллагеновые, эластические и ретикулярные волокна. 2.2. Развитие центральной нервной системы и органов чувств (рис. 5.2).
Рис. 5.2. Последовательные стадии (А—В) развития головного мозга зародыша позвоночных: вм — воронка мозга; вме — центральная мозговая складка; мп — мостовой изгиб; зп — затылочный изгиб; нкк—нервнокишечный канал; пп — нейропор; ими — первичный мозговой пузырь;.х — хорда; тн — теменной изгиб.
Нервная трубка зародышей всех позвоночных вскоре после своего замыкания состоит из более широкого переднего и более узкого заднего отдела. Расширенный передний отдел называют первичным мозговым, пузырем (первичным головным мозгом — archencephalon). Первичный головной мозг открывается наружу нейропором, а задний отдел посредством нервно-кишечного канала связан с задним отделом гастроцеля (в области дорсальной губы бластопора). Нейропор и нервно-кишечный канал впоследствии зарастают. Нервная трубка по средней линии подстилается хордой, которая доходит до задней границы первичного головного мозга. Последний подстилается тканью, происшедшей из прехордальной пластинки. Как правило, задняя граница первичного головного мозга также отмечена резкой складкой вентральной стенки нервной трубки (вентральная мозговая складка), спереди от которой вентральная стенка первичного мозгового пузыря образует воронкообразный выступ (infundibulum), или воронку мозга. Вентральная мозговая складка и воронка формируют характерный для всех позвоночных теменной, или среднемозговой, изгиб. В дальнейшем передняя часть нервной трубки дифференцируется на три мозговых пузыря: передний (prosencephalon), расположенный спереди от вентральной складки, средний (mеsencephalon), находящийся над этой складкой, и задний (rhombencephalon), без резкой границы переходящий в спинной мозг. У зародышей высших позвоночных уже на стадии трех мозговых пузырей при взгляде сверху отчетливо видны боковые выступы переднего мозгового пузыря, впоследствии дающие начало глазным зачаткам. Позже передний мозговой пузырь подразделяется на два отдела: передний (telencephalon) и промежуточный (diencephalon) мозг. Из боковых стенок последнего в дальнейшем развиваются глазные зачатки. Средний мозговой пузырь в дальнейшем не расчленяется, а первичный задний мозговой пузырь подразделяется на задний (metencephalon) и продолговатый (myelencephalon) мозг, переходящий без резкой границы в спинной мозг. У низших позвоночных эти отделы мозга лежат примерно в одной плоскости, а у высших — головной мозг вскоре после формирования названных отделов образует новые резкие изгибы: затылочный и мостовой. Затылочный изгиб находится на месте перехода спинного мозга в продолговатый и направлен в ту же сторону, что и теменной, мостовой изгиб располагается в области заднего мозга и назван так потому, что в вентральной стенке этого мозгового пузыря впоследствии возникает варолиев мост. Этот изгиб направлен -в сторону, обратную двум другим изгибам. Все мозговые изгибы особенно хорошо выражены у высших млекопитающих и человека. 2.3. Нервный гребень и его производные. При смыкании нервной трубки клетки нервных валиков располагаются над ее дорсальной частью. Образованная ими структура называется нервным гребнем. Уже в процессе замыкания нервной трубки клетки нервного гребня выходят из состава нервных валиков и мигрируют в разных направлениях, проявляя формообразовательные потенции. Некоторые клетки нервных валиков формируют хрящи висцерального скелета, мышцы кожи и ресничного тела глаза, рыхлую соединительную ткань лица, языка и нижней челюсти, а также входят в состав аденогипофиза, паращитовидных желез и мякоти зуба. Таким образом, клетки нервного гребня проявляют способность формировать такие закладки (например, хрящи), которые в других случаях возникают из мезодермы. 3. Развитие производных мезодермы. 3.1. Осевая мезодерма. У всех позвоночных имеются осевая и боковая мезодермы, причем осевая мезодерма подразделяется на сомиты (метамеризуется; рис. 5.3).
Рис. 5.3. Развитие целомической мезодермы у зародыша человека: А – поперечный разрез зародыша, имеющего около десяти сомитов (при малом увеличении микроскопа); Б – тот же разрез при большем увеличении: 1–двухслойный амнион; 2–энтодермальный эпителий тела зародыша; 3–нервная трубка; 4–сомит без ясно выраженного деления на дерма-миотом и силеротом; 5–нефротом; 6–соматоплевра; 7–спланхноплевра; 8–целомическая полость, 9–хорда; 10–однослойный энтодермальный эпителий кишечника зародыша; 11–стенка желточного мешка.
Способ закладки и дифференцировки сомитов в разных классах хордовых неодинаков. У ланцетника сомиты формируются в виде энтероцельных выпячиваний архентерона и с самого начала содержат участок целомической полости. У большинства позвоночных сомиты сначала закладываются в виде сплошных скоплений мезодермальных клеток и лишь позже в них возникают полости путем расхождения этих клеток (рис. 5.4). В ходе дальнейшего развития сомита из его клеток образую
|
||||
|
|
Последнее изменение этой страницы: 2016-06-22; просмотров: 707; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.217.242.39 (0.013 с.) |