Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Общая физиология сенсорных системСодержание книги
Похожие статьи вашей тематики
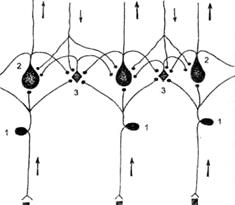
Поиск на нашем сайте ПРЕДИСЛОВИЕ ЧАСТЬ I ОБЩАЯ ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ В качестве введения в частные аспекты сенсорной физиологии, обсуждаемые в данном учебном пособии, мы рассмотрим предмет «общей физиологии сенсорных систем» - принципы, лежащие в основе сенсорных систем человека и животных. Такой подход возможен и полезен, потому что разные сенсорные системы очень схожи по своей организации и функции, по реакциям, которые они вызывают. Общий принцип работы сенсорных систем заключается в том, что деятельность любой сенсорной системы начинается с восприятия сигнала соответствующими (по модальности) сенсорными рецепторами внешней для мозга физической и химической энергии, трансформации ее в нервные импульсы и передачи их в мозг через цепи нейронов, образующих ряд уровней. Процесс передачи сенсорного сообщения сопровождается многократным преобразованием и перекодированием и завершается общим анализом и синтезом (опознаванием образа). После этого происходит выбор или разработка программы ответной реакции организма. Таким образом, с физиологической точки зрения сенсорная система выполняет такие основные функции, или операции, с сигналами как их обнаружение, различение, кодирование информации о сигнале, ее передачу и преобразование; детектирование признаков; опознание образов. Обнаружение и первичное различение сигналов обеспечивается рецепторами, а детектирование и опознание сигналов — нейронами коры больших полушарий. Передачу, преобразование и кодирование сигналов осуществляют нейроны всех слоев сенсорных систем. С точки зрения теории информации, для любых устройств, в которых происходит переработка информации, необходима быстрота передачи информации, точность ее передачи (помехоустойчивость), а также возможность кодирования и декодирования информации. В живых системах для получения такого комплекса положительных эффектов возникла система передачи информации двойничным кодом (да — нет), в которой в качестве слова «да» используется наличие потенциала действия, а в качестве «нет» — его отсутствие. Таким образом, в общем виде работа анализатора заключается в том, чтобы в рецепторах закодировать информацию о сигнале внешней или внутренней среды, а в нейронах мозга провести декодирование информации и превращение ее в факт осознаваемого (ощущение). ►Понятие о сенсорных системах. Сенсорные системы мозга (или, по И.П. Павлову, анализаторы) — это совокупность сенсорных рецепторов, специализированных вспомогательных аппаратов, нервных волокон и многочисленных нейронов мозга, которые участвуют в обработке информации о сигналах внешнего или внутреннего мира, на основе которой формируются ощущения и восприятия — основа представления о мире. Все анализаторы, по И.П. Павлову, состоят из трех основных отделов: периферического, проводникового и центрального, или коркового. В периферическом отделе с помощью сенсорных рецепторов происходит обнаружение сигналов внешнего и внутреннего мира, их первичное различение и превращение в электрический процесс. В проводниковом отделе осуществляется последовательная обработка сенсорной информации и проведение ее в высшие отделы мозга. В центральном, или корковом, отделе совершается окончательная обработка сенсорной информации и формируется вначале ощущение (т.е. представление об отдельных свойствах сенсорного сигнала, или субъективный образ сигнала), а затем восприятие (перцепция), т.е. целостное, интегральное отражение отдельных предметов или явлений внешнего мира. Именно восприятие составляет основу всей интеллектуальной деятельности человека, т.е. мышления. Выделяют такие сенсорные системы как зрительная, слуховая, вестибулярная, соматическая (в том числе тактильная, температурная и ноцицептивная, или болевая), проприоцептивная, вкусовая, обонятельная, висцеральная (интероцептивная). Итого — 10 систем. Благодаря сенсорным системам у человека формируются соответственно 10 видов ощущений, или чувств, из которых один вид ощущений (возникающий на основе интероцептивной информации) — не всегда имеет четкое осознание. В зрительной, слуховой, вестибулярной, соматической, вкусовой и обонятельной сенсорных системах периферический отдел устроен достаточно сложно, т.е. включает в себя множество морфологических структур, способствующих детекции сигнала. Для обозначения сложно устроенных периферических отделов введено такое понятие как органы чувств (огgапа sепsоrа). К ним относятся: глаз (орган зрения), ухо (орган слуха), вестибулярный аппарат (орган гравитации), кожа (орган осязания), вкусовые сосочки языка (орган вкуса) и нос (орган обоняния). В норме все сенсорные системы осуществляют свою деятельность не изолированно, а в тесном взаимодействии друг с другом. Так, функция зрительной системы изменяется под действием звукового раздражителя. При этом улучшается способность различать светлые объекты на темном фоне. Освещение глаз делает слышимые звуки более громкими и т.д. Такое сотрудничество обеспечивает одновременное восприятие различными рецепторными аппаратами сложных комплексных раздражителей, падающих на организм в обычных условиях существования. Кроме того, взаимосвязь анализаторов очень важна и в случае утраты одного из них. Так, например, отсутствие зрения компенсируется обострением слуховой и осязательной чувствительности, что позволяет слепым ходить без провожатых, «читать» с помощью пальцев рельефный текст. ►Классификация раздражителей. Раздражитель – это любое физико-химическое изменение среды (как внешней, так и внутренней), приводящее к сдвигам метаболизма. Именно раздражитель является пусковым сигналом для начала работы анализаторов. Все сигналы (раздражители) внешней и внутренней среды, которые воспринимаются сенсорными системами мозга, различаются по модальности, т.е. по той форме энергии, которая свойственна каждому из них. Раздражители бывают механические, химические, тепловые, осмотические, световые, электрические. Они передаются с помощью различных форм энергии. Так, например, свет — фотонами, химические раздражители — молекулами и ионами, механические — посредством механической формы энергии. Все раздражители независимо от их модальности подразделяются на адекватные и неадекватные. Адекватность раздражителя проявляется в том, что его пороговая интенсивность значительно ниже по сравнению с неадекватными раздражителями, например, воздействие светового и механического стимулов на рецепторы глаза. Ощущение света возникает у человека, когда минимальная интенсивность светового раздражителя составляет всего 10-17— 10-18 Вт. Но ощущение вспышки можно вызвать и при механическом воздействии на глаз (это явление называют механическим фосфеном). В этом случае мощность раздражителя должна быть не менее 10-4 Вт. Этот пример демонстрирует разницу между адекватным и неадекватным раздражителем. ГЛАВА 1 ОБЩАЯ ФИЗИОЛОГИЯ РЕЦЕПТОРОВ СЕНСОРНЫХ СИСТЕМ ►Понятие о сенсорных рецепторах. Основным компонентом периферического отдела сенсорных систем является рецептор. Он представляет собой высокоспециализированную структуру (у первичночувствующих рецепторов — это видоизмененный дендрит афферентного нейрона, у вторичночувствующих — это сенсорная рецепторная клетка), которая способна воспринимать действие адекватного раздражителя внешней или внутренней среды и трансформировать его энергию в конечном итоге в потенциалы действия — специфическую активность нервной системы. Здесь следует напомнить, что понятие «рецептор» (от лат. гесерio, гесерtum — брать, принимать) в физиологии используется в двухзначениях. Во-первых, для обозначения специфических белков клеточной мембраны или цитозоля, которые предназначены для детекции гормонов, медиаторов и других биологически активных веществ. Такие рецепторы принято называть мембранными, клеточными, или гормональными (например, альфа-адренорецепторы). Во-вторых, для обозначения рецепторов как компонентов сенсорной системы. Эти рецепторы часто называют сенсорными рецепторами, или сенсорными рецепторными клетками. ►Классификация рецепторов. В зависимости от того, из внутренней или внешней среды воспринимаются раздражения, все сенсорные рецепторы подразделяют на экстероцепторы и интерорецепторы. Экстерорецепторы воспринимают сигналы внешней среды. К ним относят фоторецепторы сетчатки глаза, фонорецепторы кортиевого органа, вестибулорецепторы полукружных каналов и мешочков преддверия, тактильные, температурные и болевые рецепторы кожи и слизистых оболочек, вкусовые рецепторы языка, обонятельные рецепторы носа. Среди интерорецепторов различают висцерорецепторы, предназначенные для детекции изменений внутренней среды, и проприрецепторы (рецепторы мышц и суставов, т.е. опорно-двигательного аппарата). Висцерорецепторы представляют собой различные хемо-, механо-, термо-, барорецепторы внутренних органов и кровеносных сосудов, а также ноцицепторы. По характеру контакта со средой экстерорецепторы делятся на дистантные, получающие информацию на расстоянии от источника раздражения (зрительные, слуховые и обонятельные) и контактные — возбуждающиеся при непосредственном соприкосновении с раздражителем (вкусовые, тактильные). В зависимости от вида модальности воспринимаемого раздражителя, т.е. от природы раздражителя, на который рецепторы оптимально настроены, сенсорные рецепторы делятся на 6 основных групп: механорецепторы, терморецепторы, хеморецепторы, фонорецепторы, ноцицепторы и электрорецепторы (последние выявлены только у некоторых рыб и амфибий). Механорецепторы приспособлены к восприятию механической энергии раздражающего стимула. Они входят в состав соматической (тактильной), скелетно-мышечной, слуховой, вестибулярной и висцеральной сенсорных систем, а также (у рыб и амфибий) сенсорной системы боковой линии. Терморецепторы воспринимают температурные раздражения, т.е. интенсивность движения молекул, и входят в состав температурной сенсорной системы. Они представлены тепловыми и холодовыми рецепторами кожи, внутренних органов и термочувствительными нейронами гипоталамуса. Хеморецепторы чувствительны к действию различных химических веществ и входят в состав вкусовой, обонятельной и висцеральной сенсорных систем. Фоторецепторы воспринимают световую энергию и составляют основу зрительной сенсорной системы. Болевые (ноцицептивные) рецепторы воспринимают болевые раздражения, в том числе механоноцицепторы — действие чрезмерных механических стимулов, хемоноцицепторы — действие специфических болевых медиаторов; они являются начальным компонентом ноцицептивной сенсорной системы. Электрорецепторы, выявленные в составе боковой линии ряда рыб и амфибий, чувствительны к действию электромагнитных колебаний. Следует подчеркнуть, что в процессе эволюции отбирались те рецепторы и соответствующие им сенсорные системы, которые обеспечивали каждый организм достаточным количеством информации, необходимой для его нормального существования и адаптации во внешней среде. В этом плане можно привести образно сказанную фразу (А.Д. Ноздрачев и соавт., 1991): «У человека не обнаружены электрорецепторы, существующие у рыб; нет рецепторов, воспринимающих прямое инфракрасное излучение, как у гремучей змеи; глаз человека не воспринимает поляризацию света, как глаза некоторых насекомых, его ухо не ощущает ультразвуковых колебаний, как слуховой аппарат летучих мышей и многих ночных млекопитающих». Но, в целом, имеющиеся у человека сенсорные системы позволяют ему успешнее других представителей животного мира осваивать Землю. Кроме представленных двух классификаций важным является деление всех сенсорных рецепторов в зависимости от их структуры и взаимоотношения с афферентным сенсорным нейроном на два больших класса — первичночувствующие (первичные) и вторичночувствующие (вторичные) рецепторы. Это определяет избирательную чувствительность рецептора к адекватным раздражителям (у вторичночувствующих она намного больше, чем у первичночувствующих), а также последовательность трансформации энергии внешнего сигнала в потенциал действия нейрона. К первичным сенсорным рецепторам относят те рецепторы, которые представляют собой видоизмененное, специализированное окончание дендрита афферентного нейрона. Это означает, что афферентный нейрон непосредственно (т.е. первично) взаимодействует с внешним стимулом. К первичночувствующим рецепторам относятся отдельные виды механорецепторов (свободные нервные окончания кожи и внутренних органов), холодовые и тепловые терморецепторы, ноцицепторы, мышечные веретена, сухожильные рецепторы, суставные рецепторы, обонятельные рецепторы. Вторичные рецепторы — это специально приспособленные для восприятия внешнего сигнала клетки ненервного происхождения, которые при своем возбуждении в ответ на действие адекватного раздражителя передают сигнал (как правило, с выделением медиатора из синапса) на дендрит афферентного нейрона. Следовательно, в этом случае нейрон воспринимает раздражитель косвенно, опосредовано (вторично) за счет возбуждения сенсорной клетки-рецептора (рецептирующей клетки). К вторичночувствующим рецепторам относятся многие виды механорецепторов кожи (например, тельца Пачини, диски Меркеля, клетки Мейсснера), фоторецепторы, фонорецепторы, вестибулорецепторы, вкусовые рецепторы, а также электрорецепторы рыб и амфибий. ►Адаптация сенсорных рецепторов. Сенсорные рецепторы способны к адаптации, которая состоит в том, что при постоянном воздействии стимула на сенсорный рецептор его возбуждение слабеет, т.е. снижается величина рецепторного потенциала, а также частота генерации потенциалов действия афферентным нейроном. Подобное явление наблюдается и при гормонрецепторном взаимодействии. В этом случае оно называется десенситизация и связано с нарушениями передачи сигнала «вниз по течению». Адаптация сенсорных рецепторов имеет еще более сложную природу. С одной стороны, она зависит от процессов, которые происходят на этапе взаимодействия сенсорного стимула с «активным центром» сенсорного рецептора (по сути, это явление десенситизации). С другой стороны, адаптация рецепторов связана с потоком импульсов, приходящим к сенсорному рецептору по эфферентным волокнам от вышележащих нейронов мозга (в том числе от нейронов ретикулярной формации), т.е. является активным процессом. В определенной степени, адаптация может быть обусловлена свойствами и состоянием вспомогательных структур периферического отела сенсорной системы. В целом, адаптация проявляется в снижении абсолютной и повышении дифференциальной чувствительности сенсорной системы. Скорость адаптации для разных рецептов различна: наибольшая для тактильных рецепторов, а наименьшая – для вестибулярных и проприорецепторов. Благодаря высокой скорости адаптации тактильных рецепторов мы быстро перестаем ощущать надетые очки, часы или одежду, а благодаря низкой скорости адаптации мышечных рецепторов можем совершать высоко координированные и четкие движения. ►Основные этапы преобразования энергии внешнего стимула в рецепторный потенциал (механизмы возбуждения сенсорных рецепторов). При всем многообразии морфофункциональных особенностей сенсорных рецепторов общую схему этого процесса можно представить в виде некоторой обобщенной схемы. В первичных рецепторах условно можно выделить пять основных этапов трансдукции сенсорного сигнала: 1) взаимодействие воспринимаемого стимула с «активным» участком сенсорного рецептора; 2) изменение ионной проницаемости мембраны; 3) уменьшение уровня мембранного потенциала сенсорно го рецептора, т.е. генерация рецепторного потенциала, уровень которого зависит от величины воспринимаемого стимула; 4) генерация потенциалов действия или увеличение частоты генерации спонтанных потенциалов действия в соме афферентного нейрона (аксонном холмике); 5) распространение потенциалов действия по аксону до второго афферентного нейрона данной сенсорной системы. Во вторичночувствующих сенсорных клетках первые три этапа идут по такой же схеме; затем добавляется еще два промежуточных этапа — 4а) выделение под влиянием рецепторного потенциала квантов медиатора (например, ацетилхолина) в синапсе рецепторной клетки; 5а) ответ дендрита афферентного нейрона на выделение медиатора генерацией возбуждающего постсинаптического потенциала, или генераторного потенциала. Остальные два этапа (4 и 5) идут так же, как и в первичночувствующих рецепторах. Единственным исключением из этого правила является цепь событий в зрительной сенсорной системе, в которой в ответ на действие света фоторецепторная клетка повышает свой мембранный потенциал, в результате чего в ней снижается продукция тормозного медиатора, что приводит к конечном итоге к возбуждению биполярного нейрона, который в свою очередь возбуждает ганглиозную клетку. ГЛАВА 2 СЕНСОРНЫХ СИСТЕМ ►Пути проведения информации от рецепторов к высшим отделам мозга. Как мы уже знаем, афферентные нейроны — это первые нейроны, которые участвуют в обработке сенсорной информации. Как правило, они локализованы в ганглиях (спинномозговые ганглии, ганглии головы и шеи, например, вестибулярный ганглий, спиральный ганглий и т. п.). Исключением являются фоторецепторы — их афферентные нейроны (ганглиозные клетки) лежат непосредственно на сетчатке. Нервные импульсы по аксонам афферентных нейронов направляются в кору больших полушарий через ряд сенсорных центров, в каждом из которых осуществляется необходимый анализ соответствующей сенсорной информации, а также интеграция этой информации с другими типами информации. Весь восходящий поток информации от периферического отдела каждой сенсорной системы (или все центральные пути) можно разделить на четыре отдельных потока: - специфический; - ассоциативный; - неспецифический; - передаточный. Одновременно в конструкции мозга предусмотрен нисходящий, эфферентный путь, который регулирует пропускную способность центральных путей и тем самым контролирует объем и качество поступающей к коре больших полушарий сенсорной информации. Благодаря взаимосвязанной деятельности специфических, ассоциативных и неспецифических образований мозга, а также эфферентных нисходящих путей обеспечивается формирование поведенческого статуса всего организма, адекватного текущему моменту. Рассмотрим особенности проведения информации по каждому из указанных путей. Специфический сенсорный путь предназначен для оценки физических параметров сенсорных стимулов, воспринятых рецепторами одного типа. Благодаря такой строгой мономодальности этот путь создает условия для формирования конкретного ощущения. Специфический сенсорный путь представлен сенсорными нейронами, передающими импульсацию от нейронов спинного мозга или ствола мозга к нейронам специфических ядер таламуса (например, тонкий и клиновидный пучки спинного мозга). Исключением является обонятельный анализатор, у которого информация от нейронов обонятельной луковицы передается к нейронам обонятельного мозга минуя специфические ядра таламуса. От проекционных ядер таламуса, который нередко называют общим сенсорным коллектором, информация поступает в соответствующие первичные проекционные зоны коры. От них она поступает ко вторичным проекционным зонам коры, от которых она доходит к передним и задним ассоциативным зонам коры (по И. П. Павлову, проекционные зоны коры — это ядерные зоны анализатора, ассоциативные — это его рассеянные зоны). В первичных проекционных зонах коры расположены мономодальные специфические нейроны, реагирующие на определенные признаки, например, на появление красного света, на движение точки вправо (главным образом, это нейроны 4-го, т.е. внутреннего зернистого слоя коры). Поражение первичных зон соответствующей сенсорной системы вызывает нарушение способности восприятия и тонкой дифференцировки раздражителей (клинически это выражается, например, аносмией или агевзией). Во вторичных проекционных зонах, прилегающих к первичным зонам, содержатся нейроны, которые возбуждаются на сложные признаки (т.е. они являются детекторами сложных признаков), но сохраняющие свою модальность (в основном, это нейроны 2-го, или наружного зернистого слоя, и 3-го слоя, или слоя пирамидных клеток коры). Так, например, для зрительной сенсорной системы первичная проекционная зона в основном представлена полем 17 по Бродману, а вторичная — полями 18 и 19 (затылочная доля); для слуховой — соответственно полями 41 и 42 и полями 21 и 22 (височная доля), для соматической сенсорной системы — полями 1 и 3 и полями 2 и 5 (теменная доля), для проприоцептивной — полем 4 и полем 6 (лобная доля); для вкусовой — полем 43 и полями 28 и 34 (теменная и височная доли). Ассоциативные зоны коры представляют собой скопление нейронов в лобной доле коры (передняя ассоциативная зона, или таламолобная ассоциативная кора), а также на границах теменной, височной и затылочной долей (задняя ассоциативная область, или таламотеменная ассоциативная кора), которые преимущественно сконцентрированы во втором и третьем слоях коры. Нейроны ассоциативных зон коры — это преимущественно полимодальные нейроны, получающие сенсорную информацию от мономодальных нейронов (зрительных, слуховых) проекционных зон коры, а также от нейронов ассоциативных ядер таламуса. С участием нейронов ассоциативной коры осуществляется детекция обобщенных признаков сенсорных стимулов, формирование «сенсорной модели мира», а также гностические процессы — узнавание, опознание образа, т.е. акцепция. При повреждении этих зон нередко возникают агнозии, т.е. нарушение узнавания (слуховая, цветовая, тактильная и др.), а также нарушение речи, в том числе моторная афазия при нарушении передней ассоциативной зоны и сенсорная афазия при повреждении задней ассоциативной зоны коры. Для осуществления анализа сенсорной информации в ассоциативных зонах коры используются нейроны с простыми, сложными и «сверхсложными» рецептивными полями, ансамбли нейронов, выделяемые «подобразы», а также многочисленные гностические («бабушкины») нейроны, узнающие определенный предмет внешнего мира (более подробно о нейронах-детекторах изложено ниже). При этом нейроны сгруппированы в виде вертикальных колонок (от одной тысячи до десяти тысяч в колонке), благодаря которым детектируются все возможные признаки сенсорного сигнала. С этой точки зрения любую сенсорную область коры можно представить как совокупность большого числа колонок, специализирующихся на процессах обработки определенного сенсорного сигнала или полимодального сигнала. Следует отметить, что колончатый принцип обработки сенсорной информации используется не только в ассоциативных зонах коры, но и в проекционных ее зонах; различия касаются набора нейронов и их способности проводить анализ сенсорной информации. Таким образом, в проекционных зонах происходит декодирование информации, на основании чего возникает представление о модальности сигнала, о его силе и качестве, а в ассоциативных участках коры — определение «что это такое?», т.е. акцепция сигнала. Все эти процессы совершаются с использованием механизмов памяти. Ассоциативный, или таламокортикальный, путь связан с оценкой биологической и социальной значимости сенсорных стимулов, т.е. с формированием восприятия. Для этих целей информация, идущая от сенсорных афферентных нейронов, направляется не только к проекционным ядрам таламуса, но и к ассоциативным его ядрам (вместе они составляют специфические ядра таламуса). В нейронах этих ядер происходит межсенсорная, или кроссмодальная интеграция, т.е. комплексная обработка информации, поступающей от различных сенсорных систем мозга. В таком виде она отправляется к нейронам ассоциативных зон коры больших полушарий, в том числе по таламопариетальному и таламофронтальному путям. В совокупности с информацией, поступающей сюда от проекционных зон коры, это позволяет получить комплексное представление о предметах внешнего мира. Неспецифический, или мультимодальный, путь предназначен для поддержания общего уровня возбудимости мозговых структур (главным образом, нейронов коры), участвующих в обработке сенсорной информации, которая поступает в эти участки, как мы уже это знаем, по специфическим и ассоциативным путям. Этот важный для обработки сенсорной информации путь представляет собой коллатеральные ответвления (коллатерали) от центрального потока к нейронам ретикулярной формации ствола мозга и неспецифических ядер таламуса. Любая сенсорная информация, независимо от ее модальности, за счет дивергенции возбуждает эти нейроны, благодаря чему осуществляется их восходящее активирующее влияние на структуры сенсорной коры. Деятельность этих двух активирующих систем во многом связна с базальными ганглиями (главным образом, хвостатым ядром). Передаточный путь предназначен для использования сенсорной информации на нижних этажах мозга для реализации многочисленных безусловных рефлексов, а также для регуляции центрального потока сенсорной информации. Этот путь представляет собой коллатеральные ответвления сенсорной информации определенной модальности на различные нейроны (включая мотонейроны и вегетативные нейроны) спинного, продолговатого и среднего мозга, а также мозжечка, гипоталамуса и лимбической системы мозга. Так, например, при нанесении болевого раздражителя на кожу правой ноги возникает безусловный сгибательный рефлекс, обусловленный тем, что импульсация от тактильных и болевых рецепторов кожи на уровне спинного мозга по коллатералям поступает к альфа-мотонейронам мышц-сгибателей правой ноги, а через вставочные нейроны она достигает альфа-мотонейронов мышц левой ноги, вызывая их торможение. ►Эфферентные, или нисходящие, пути и их роль в обработке сенсорной информации. Они представлены многочисленными аксонами нейронов коры больших полушарий, таламуса и других подкорковых образований, которые оканчиваются на различных уровнях каждой сенсорной системы, включая ее периферический отдел. Общее число эфферентных нервных волокон, приходящих к рецепторам или элементам какого-либо нейронного слоя сенсорной системы, как правило, во много раз больше числа афферентных волокон, приходящих к тому же слою. Это определяет важную особенность эфферентного контроля в сенсорных системах: его широкий и диффузный характер. С помощью эфферентных путей высшими отделами мозга регулируется (уменьшается или увеличивается) восходящий поток сенсорной информации, и тем самым оптимизируется процесс обработки сенсорной информации в подкорковых структурах мозга и в коре больших полушарий. Это достигается различными способами, в том числе за счет изменения чувствительности сенсорного рецептора (например, при возбуждении гамма-мотонейронов спинного мозга возрастает чувствительность мышечного веретена к растяжению), за счет регуляции величины рецептивного поля (при темновой адаптации под влиянием эфферентных импульсов возрастает число фоторецепторов, конвергирующих на одной ганглиозной клетке), за счет регуляции адаптационных процессов в сенсорных рецепторах (чаще всего, речь идет о тормозных влияниях, уменьшающих чувствительность рецепторов и/или ограничивающих поток сенсорной информации), а также за счет вытормаживания, или избирательной фильтрации избыточной ин формации, благодаря чему к центрам в «концентрированном» виде поступает наиболее важная, наиболее содержательная и наиболее обобщенная информация. Последнее достигается тем, что от нейронов среднего мозга, таламуса и коры больших полушарий идут нисходящие тормозные пути, которые по мере необходимости тормозят проведение сенсорной информации в центр. Существование нисходящих путей, с одной стороны, подчеркивает, что спинальные, стволовые и таламические структуры сенсорных систем не просто релейные станции, а места контроля передачи афферентной импульсации с нижележащих уровней на вышележащие, а с другой стороны - позволяет говорить о важной роли обратных связей в деятельности сенсорных систем. ►Основные принципы функционирования проводникового и коркового отделов сенсорных систем. Принцип многоканальности означает наличие на каждом уровне сенсорной системы множества нейронов, связанных с множеством нейронов следующего уровня, что обеспечивает сенсорной системе точность и детальность анализа сигналов. Кроме того, дублирование каналов обработки и передачи информации является одним из путей обеспечения надежности работы сенсорных систем. Поэтому существование наряду с вышеописанными и спрямленных путей уменьшает число промежуточных звеньев переключения импульсации и коэффициент информативности передаваемого по этому каналу сигнала должен возрастать. Появление таких каналов отражает общую тенденцию к совершенствованию конструкции мозга и повышению надежности работы его сенсорных аппаратов. Принцип многоуровневости. Поканальность проведения сигнализации предполагает многоуровневый или многоэтажный характер передачи и обработки сенсорных сообщений. Чем большее число каналов в пределах данной сенсорной системы, тем большее число переключений (своего рода передаточных «станций») характерно для каждого канала. Однако такие «станции» выполняют не только функцию сенсорных реле, но и функцию обнаружения, а также служат фильтрами или центрами перекодирования. Такая многоканальность, наряду с более детальной обработкой информации, подразумевает ее замедленную передачу в высшие мозговые центры. Но в то же время это позволяет организму быстро реагировать на простые сигналы, анализируемые уже на первых уровнях сенсорной системы. Так, например, реализация ориентировочного рефлекса с участием ядер крыши среднего мозга значительно повышает шансы организма на выживание, подготавливая его к реакции на новый внезапный световой или слуховой раздражитель. Человек, имеющий нарушения в этой области мозга, не способен к быстрой ответной реакции на неожиданное раздражение. Однако при этом в каждой сенсорной системе, по-видимому, существуют афферентные каналы срочной передачи (без переключений) в проекционные зоны коры больших полушарий, благодаря чему осуществляется преднастройка высших мозговых центров к восприятию последующей информации. Принцип конвергенции и дивергенции, или принцип «сенсорной воронки». Надежность каналов связи еще более возрастает благодаря частичному взаимному перекрытию нейронов. Даже на рецепторном уровне, например, в сетчатке глаза, концевые разветвления одного и того же нейрона контактируют с несколькими нейронами более высокого уровня (дивергенция). В то же время один и тот же нейрон контактирует, получая импульсацию, сразу с несколькими нейронами предыдущего уровня (конвергенция). Следовательно, четкая линейность проведения информации по независимым каналам сенсорной системы не соблюдается. Это обусловлено наличием различного числа рецепторов и нейронов на соседних уровнях сенсорной системы. Например, у человека на сетчатке насчитывается 130 млн. фоторецепторов, а ганглиозных клеток, на которые передается сигнал от них, в 100 раз меньше («суживающаяся воронка»). На следующих уровнях формируется «расширяющаяся воронка» - число нейронов в первичной проекционной зоне в тысячи раз больше, чем ганглиозных клеток сетчатки. Физиологический смысл «суживающейся воронки» заключается в уменьшении избыточности информации, а «расширяющейся» - в обеспечении дробного и сложного анализа разных признаков сигнала. По-видимому, оба этих механизма необходимы для неискаженной передачи сведений об отдельных признаках и деталях наряду с объединением их при формировании целостного образа. Принцип обратных связей. Деятельность сенсорной системы должна рассматриваться не как процесс пассивного кодирования любого раздражения в частотно модулированную импульсную сигнализацию, а как процесс активного восприятия и обработки наиболее биологически существенной информации. Сенсорную систему необходимо рассматривать не просто как пассивный канал линии связи, а как активный «сигнализатор». Сенсорная система состоит не из лестницы релейных образований, а из аппаратов управления процессом обработки и передачи информации с нижележащих уровней на вышележащие. И каждый уровень системы должен работать минимум на основе двух входов: 1) входа информации — восходящий путь и 2) входа управления — нисходящий путь. Таким образом, центральная организация сенсорной системы в совокупности прямых и обратных связей представляется в виде системы образований из надстраивающихся нервных колец (рис. 2.1).
Рис.2.1. Схема построения нейронных переключений в сенсорной системе. Периферический чувствительный нейрон (1), передающий центростремительные импульсы как на афферентные нейроны (2) с длинным аксоном, так и на специальные переключательные нейроны (3) с коротким аксоном. Наряду с восходящими проводниками (стрелки вверх) показаны нисходящие центробежные проводники (стрелки вниз).
Наличие нисходящих связей к различным образованиям сенсорной системы говорит о том, что на их работу влияют вышерасположенные отделы многих мозговых структур. Благодаря деятельности нисходящих связей обеспечивается регуляция пороговой чувствительности и пропускной способности в сенсорных системах. Это естественно, поскольку сенсорная система может обработать за единицу времени объем информации во много раз меньший, чем тот, который может поступать на ее вход. Направленность нисходящих влияний определяется доминирующей потребностью как предпосылкой образования доминанты при использовании аппаратов памяти. В связи с этим особенно важной оказывается кортикальная регуляция, которая осуществляет три функции: 1) пусковую, 2) корригирующую и 3) поддерживающую. Значение первой из них состоит в открытии или блокировании сенсорных входов; второй — в трансформации нервной сигнализации или в обеспечении условий, при которых ранее разбалансированные сенсорные системы могут быть приведены к такому режиму деятельности, который создает необходимые условия для их наиболее полного взаимодействия и интеграции, и третий — в поддержании возбудимости глубоких структур данной сенсорной системы на уровне, оптимально приспособленном к восприятию и передаче сигнала в восходящем направлении. Имеющиеся данные свидетельствуют о тормозном значении обратных связей. Это становится понятным, если принять во внимание чрезвычайно высокую (можно сказать, критическую) чувствительность рецепторных образований сенсорных систем. Именно на этих уровнях сенсорных систем начинается процесс активного восприятия и обработки сигналов. Принцип двусторонней симметрии. Любая сенсорная система построена по принципу билатеральной симметрии. Иными словами, рецепторные аппараты, соответствующие им центральные проводники и центральные мозговые структуры парные, и одна половина зеркально повторяет другую. Однако, как правило, этот принцип проявляется лишь в относительной степени, поскольку даже первичный сенсорный путь может быть связан с обоими симметричными мозговыми полушариями. Связь с контралатеральным полушарием выражена сильнее благодаря большему числу направляющихся туда сенсорных волокон. Между симметричными отделами сенсорных систем устанавливаются горизонтальные комиссуральные связи, обеспечивающие их взаимодействие. Принцип структурно-фун |
|||
|
|
Последнее изменение этой страницы: 2016-04-18; просмотров: 777; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.102 (0.013 с.) |