Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тактильная (осязательная) сенсорная системаСодержание книги
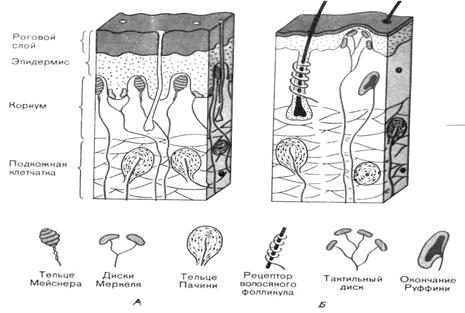
Поиск на нашем сайте Тактильная сенсорная система, наряду с проприорецептивной, зрительной и вестибулярной сенсорными системами, «поставляет» головному мозгу информацию о положении и движении тела в пространстве, о положении его отдельных частей. Кроме того, она играет важную роль в ориентации человека в окружающей среде (особенно сильно кожное осязание развивается у слепых и слепоглухонемых, позволяя тем самым таким людям избегать воздействия повреждающего агента). Благодаря тактильной сенсорной системе осуществляется контакт грудного ребенка с матерью, выполнение различных игровых, образовательных и трудовых операций, интимные отношения между мужчиной и женщиной, ощущение комфорта от носимой одежды. В принципе, такие примеры, доказывающие значение тактильной рецепции, можно перечислять еще и еще. Скажем только одно – нарушение этого вида чувствительности существенно снижает адаптационные возможности человека и лишает его многих радостей жизни. Особое место тактильная сенсорная система занимает у млекопитающих, выполняя жизненно важные функции – тактильное исследование окружающей среды, питание, звукообразование и т.д. Тактильная чувствительность резко повышена на участках тела, которые покрыты волосами. Это объясняется тем, что волосы играют роль рычажков и усиливают передачу раздражения в несколько раз, а так как 95% поверхности человеческого тела покрыто волосами, на некоторых участках едва заметными, то любое прикосновение к поверхности тела резко усиливается. ►Рецепторы осязательного анализатора. Различаютследующие основные механорецепторы – свободные неинкапсулированные нервные окончания, свободные нервные окончания волосяных фолликулов, диски Меркеля, тельца Руффини, тельца Мейснера и тельца Пачини. Строение указанных рецепторов различно, они распределены неравномерно и локализуются на разной глубине кожи. Первые два типа рецепторов относятся к первичным (являются окончаниями дендрита чувствительного нейрона), остальные – ко вторичным (представляют собой инкапсулированные специализированные клетки, трансформирующие механическое воздействие в рецепторный потенциал, который передается на дендрит чувствительного нейрона). Рассмотрим отдельные виды рецепторов (рис. 4.1).
Рис. 4.1. Схема строения и положения механорецепторов в коже, на не покрытых волосами (А) и волосистых (Б) участках кожи.
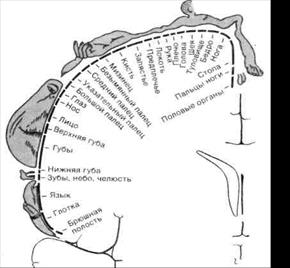
Свободные неинкапсулированные нервные окончания являются самыми распространенными рецепторами кожи. Они находятся в основном в сосочковом слое дермы – обычно идут вдоль мелких сосудов и представляют собой разветвления дендрита афферентных нейронов. Первоначально их считали рецепторами боли, но в настоящее время их рассматривают как полимодальные рецепторы, отвечающие и на болевые, и на температурные, и на механические стимулы. Это медленно адаптирующиеся рецепторы: продолжают отвечать весь период времени, пока действует стимул. Свободные нервные окончания волосяных фолликулов также представляют собой разветвления дендрита афферентного нейрона, которые оплетают волосяную сумку. Обычно фолликул получает волокна от нескольких чувствительных нейронов, но в то же время один и тот же дендрит чувствительного нейрона иннервирует несколько волосяных фолликулов. Волос, как было уже отмечено выше, служит рычагом, усиливающим раздражение нервных окончаний, что и объясняет высокую чувствительность волос к прикосновению. Указанные рецепторы реагируют преимущественно на легкое прикосновение и осуществляют пространственное и временное тактильное различение. Диски (клеточные комплексы) Меркеля представляют собой видоизмененные эпителиальные клетки, с которыми образуют синапсы дендриты афферентных нейронов. Находятся в базальном слое эпидермиса и частично в сосочковом слое дермы в виде небольших скоплений. Особенно много дисков Меркеля в участках кожи с высокой чувствительностью, т.е. в голой коже пальцев рук и на губах. В покрытой волосами коже они лежат в особых колоколообразных тельцах, выдающихся над окружающей кожной поверхностью (в каждом таком образовании, называемом тактильным тельцем Пинкуса-Игго, находится 30-50 клеток Меркеля, связанных с афферентными нервными волокнами). При высоте около 0,1 мм и диаметре 0,2-0,4 мм эти тельца едва различимы невооруженным глазом. Поскольку диски Меркеля медленно адаптирующиеся рецепторы, то их рассматривают в качестве пропорционального датчика, т.е. генерация рецепторного потенциала в них тем активнее, чем сильнее действует раздражитель. Эти рецепторы рассматривают как рецепторы давления, или силы, поскольку они воспринимают изменение силы механического воздействия. Тельца (окончания) Руффини располагаются в волосистой части кожи – в глубоких слоях эпидермиса и в сосочковом слое дермы. Представляют собой веретеновидной формы капсулу, образованную плотно переплетенными коллагеновыми волокнами. Внутри такой капсулы содержится жидкость, в которую погружены разветвления дендрита чувствительного нейрона. Как и диски Меркеля, являются рецепторами давления, или силы. Тельца Мейснера (тактильное мейснеровое тельце) представляют собой капсулу конусовидной или овальной формы. Капсула ориентирована перпендикулярно поверхности кожи. Ее стенки образованы многочисленными пластинчатыми клетками, между которыми параллельно поверхности кожи располагаются терминали дендрита чувствительного нейрона. Эти рецепторы находятся в сосочках собственно кориума, в участках, лишенных волос (кожа ладоней, подошв, пальцев рук и ног, а также губы, сосочки молочных желез и половые органы). В покрытой волосами коже их меньше и здесь они принимают форму рецепторов волосяных фолликулов. Реагируют на скорость изменения силы, т.е. являются датчиком скорости. Тельца Пачини (пластинчатые тельца, тельца Фатера-Пачини) – наиболее крупные и наиболее распространенные в организме рецепторы, а потому они являются наиболее изученными рецепторами кожи. Располагаются в гиподерме и частично в глубоких слоях дермы. Кроме того, они обнаружены и в сухожилиях и фасциях мышц, в надкостнице, в суставных сумках. Тельце Пачини напоминает луковицу, состоит из наружной капсулы, внутренней колбы и заключенного в нее дендрита афферентного нейрона. Пространство между наружной капсулой и внутренней колбой, а также внутри колбы заполнено жидкостью (ликвором). Возбуждение этих рецепторов происходит при кратковременном (в течение 0,1 мс) смещении капсулы на 0,5 мкм. Тельца Пачини считаются датчиком ускорения. Благодаря своим свойствам они воспринимают самую незначительную деформацию кожи при соприкосновении с различными предметами и почвой, т.е. с какой скоростью смещается кожа. Таким образом, большинство тактильных рецепторов локализовано, как правило, в глубоких слоях эпидермиса и сосочковом слое кориума. Всех их можно разделить на три главных типа: - рецепторы давления (датчики силы), которые ведут себя как пропорциональные датчики, т.е. генерация рецепторного потенциала в них тем активнее, чем сильнее действует раздражитель. Поэтому их еще называют пропорциональными рецепторами. Это свободные неинкапсулированные нервные окончания, диски Меркля, тельца Руффини; - рецепторы прикосновения (датчики скорости) реагируют на скорость изменения силы, т.е. скорость вдавления стимула, поэтому осуществляют пространственное и временное тактильное различение. Это тельца Мейсмера и рецепторы волосяных фолликулов; - рецепторы вибрации (датчики ускорения) – тельца Пачини - реагируют на изменение скорости механического воздействия. Однако при этом необходимо подчеркнуть, что механические стимулы, обычно действующие на кожу, одновременно возбуждают в той или иной степени несколько типов механорецепторов в зависимости от характера стимула. Поэтому возникающие ощущения нельзя приписать рецепторам одного типа. Соответственно в повседневной жизни трудно определить различия между ощущениями давления и прикосновения. Принцип работы механорецепторов. Независимо от того, что одни рецепторы реагируют на изменение силы, вторые – на скорость изменения этой силы, а третьи – на вторую производную изменения силы, во всех случаях принцип работы рецептора состоит в том, что под влиянием механического стимула в мембране рецептора изменяется ионная проницаемость, что приводит к появлению рецепторного потенциала. Он вызывает выделение медиатора, что сопровождается появлением генераторного потенциала в дендритах чувствительного нейрона, благодаря которому изменяется частота генерации потенциалов действия. Различия в восприятии механических стимулов определяются скоростью адаптации тактильных рецепторов. Так, быстро адаптирующиеся рецепторы являются датчиками скорости, или ускорения, а медленно адаптирующиеся – датчиками изменения силы. В то же время скорость адаптации определяется структурой рецепторов – наличие сложно организованной капсулы рецептора повышает скорость его адаптации (укорачивает рецепторный потенциал), поскольку такая капсула хорошо проводит быстрые и гасит медленные изменения давления. Адаптация механорецепторов кожи имеет большое значение – благодаря этому свойству рецепторов мы перестаем ощущать постоянное давление одежды, привыкаем носить на роговице глаз контактные линзы, а на носу – очки и т.д. ►Проведение осязательной информации в центральный отдел. Сенсорная информация от механорецепторов кожи передается в кору больших полушарий по нервным волокнам тонкого и клиновидного пучков, образующих лемнисковую систему. Тонкий пучок, или пучок Голля, несет информацию от рецепторов нижних конечностей и нижней части туловища, а клиновидный пучок, или Бурдаха, - от рецепторов верхних конечностей и верхней части туловища. Оба пути состоят из афферентных нейронов (они находятся в спинномозговых узлах), аксоны которых вступают в задние рога спинного мозга и, не прерываясь, направляются в составе задних столбов до тонкого (ядро Голля) и клиновидного (ядро Бурдаха) ядер продолговатого мозга. Аксоны нейронов этих ядер полностью перекрещиваются на уровне олив, образуя медиальный лемниск (петлю), и, поднимаясь дальше, заканчиваются в специфических ядрах таламуса, которые из-за своего анатомического положения называют вентробазальным комплексом. Нейроны специфических ядер таламуса посылают свои аксоны в соматосенсорные зоны коры больших полушарий (SI и SII). Поражение задних столбов клинически проявляется в потере мышечно-суставного чувства, вибрационной и тактильной чувствительности на стороне поражения при сохранении болевой и температурной чувствительности. Лемнисковая система проводит точную (по силе и месту воздействия) и сложную (о давлении, прикосновении, вибрации и движении в суставах) информацию с большой скоростью (до 80 км/с). Для всех составляющих лемнисковой системы (задних столбов спинного мозга, тонкого и клиновидного ядер, таламических ядер и корковых областей) выявлена четкая топографическая организация проекции кожи, т.е. каждый участок кожи контрлатеральной части тела занимает определенную зону, площадь которой зависит от функционального значения этой части тела. Часть импульсов от рецепторов давления кожи и слизистой передается в кору больших полушарий и по вентральному спиноталамическому тракту, который проходит в составе передних столбов спинного мозга. Однако в спиноталамической системе на уровне таламических ядер правильная соматотопическая организация представительства поверхности тела отсутствует. Особое место в передаче сенсорной информации от механорецепторов отводится V паре черепно-мозговых нервов - тройничному нерву, который в своих трех ветвях содержит афференты, идущие от лица и полости рта. Он иннервирует кожу, зубы, слизистую полости рта, язык и роговицу. Ко времени рождения тройничная система уже хорошо развита и проводит сенсорные сигналы, которые запускают пищевое поведение. Именно она обеспечивает первое сенсорное знакомство новорожденного с окружающим миром. ►Корковый уровень анализа тактильной информации. Информация от нейронов специфических ядер таламуса первоначально поступает в две проекционные соматосенсорные зоны коры больших полушарий (SI и SII). В частности, информация от нейронов вентробазального комплекса направляется (контрлатерально) в первую проекционную зону, которая у приматов и человека находится в постцентральной извилине (SI). От нейронов задней группы ядер таламуса информация преимущественно поступает (контрлатерально и ипсилатерально) во вторую проекционную соматосенсорную зону коры (SII), которая расположена в области сильвиевой (латеральной) борозды (рядом со слуховой зоной). От этих двух проекционных соматосенсорных зон информация поступает в передние и задние ассоциативные зоны коры. Первая проекционная соматосенсорная область, локализованная в постцентральной извилине (первичные зоны — 1-е и 3-е поля по Бродману, вторичные зоны — 2-е и 5-е поля), является, по сути, местом окончания лемнискового пути и ядром тактильного анализатора. Она отличается от других областей коры очень высокой степенью топографической организации (проекции различных областей поверхности тела на соматосенсорную кору осуществляются по принципу "точка в точку"). Такое явление называют соматотопией, или топографическим представительством. О наличии соматотопии свидетельствует тот факт, что при раздражении поверхности кожи короткими точечными прикосновениями первичный ответ в соматосенсорной коре локализуется в строго ограниченном участке. Показано, что размеры представительства соответствующих участков поверхности кожи у человека и приматов в постцентральной извилине связаны не с величиной поверхности их тела, а зависят от биологической значимости информации, воспринимающейся тем или иным участком кожи. Например, у человека представительство губ, лица и кистей рук в этой зоне по площади намного больше, чем представительство туловища и нижних конечностей. Это дает специфический рисунок сенсорного гомункулюса — отражения тела человека в постцентральной извилине (рис. 4.2). Характер этого рисунка свидетельствует о высокой чувствительности и тонком различении для этих частей тела, а также их биологической значимости. Так, при осмотре какого-либо предмета, когда нам надо выяснить форму, наличие шероховатостей и т.д., мы поглаживаем этот предмет, т.е. касаемся его поверхности кожей нашей ладони. Когда кора в области SI разрушена повреждением или удалена с терапевтической целью, возникает дефицит восприятия. Стимуляция кожи еще может быть воспринята как таковая, но способность локализовать ее и распознать пространственные детали раздражителя нарушается. Так, например, больные не могли на ощупь отличить овал от параллелепипеда (в клинике это нарушение носит название астерогнозис). Степень дефицита зависит от размеров поврежденного участка коры. Здесь снова выступает соматотопическая организация. Однако надо сказать, что после достаточно долгого промежутка времени подобные нарушения становятся слабее. Такое улучшение, по-видимому, обусловлено способностью других областей коры (например, 5-го поля теменной коры, соседнего с SI) принять на себя функции SI.
Рис.4.2. Схема чувствительного гомункулюса (соматотопическая организация соматосенсорной коры SI человека). Разрез полушарий (на уровне постцентральной извилины) во фронтальной плоскости. Обозначения показывают пространственное представительство поверхности тела в коре, установленное на основании локальной стимуляции мозга бодрствующих больных.
Анализ информации от тактильных рецепторов в первой соматосенсорной зоне осуществляется нейронами, объединенными в вертикальные колонки, которые можно рассматривать как своеобразные функциональные единицы, или блоки коры. Каждая такая колонка, получая информацию от рецепторов одной и той же модальности, находящихся на одном и том же рецептивном поле кожи, проводит этот анализ с участием специализированных нейронов, число которых в колонке достигает 105. Каждый из этих нейронов «настроен» на определенный признак, наличие которого в поступающей информации вызывает возбуждение соответствующего нейрона. Благодаря деятельности колонок мозг получает информацию обо всех свойствах стимула, воздействующего на соответствующий участок кожи. Во вторую соматосенсорную зону коры больших полушарий (SII), расположенную в области сильвиевой борозды вблизи от слуховой зоны (40-е и 51-е поля), поступают импульсы от тактильных рецепторов кожи «своей» и противоположной стороны. Эта зона содержит точное и детальное представительство поверхности тела, как и первая соматосенсорная зона, с тем различием, что проекции обеих половин тела во второй соматосенсорной зоне полностью перекрываются, благодаря чему происходит объединение и сравнение информации поступающей от правой и левой половины тела, т.е. имеет место билатеральное соматотопическое представительство. Как полагают, SII специально играет роль в сенсорной и моторной координации активности на двух сторонах тела (например, хватание или ощупывание обеим руками). Не исключено, что вторая соматосенсорная зона может, кроме того, осуществлять контроль над афферентной передачей сигналов в таламических ядрах. От первичных и вторичных проекционных зон коры информация от тактильных рецепторов поступает в передние (фронтальные) и задние ассоциативные зоны коры, благодаря которым завершается процесс восприятия, т.е. происходит опознание образа (акцепция сигнала). Это реализуется с участием специальных нейронов («бабушкиных» нейронов), проходящих «обучение» в процессе индивидуального развития человека. В целом роль соматосенсорной зоны коры состоит в интегральной оценке соматосенсорных сигналов, во включении их в сферу сознания, полисенсорный синтез и в сенсорное обеспечение выработки новых двигательных навыков. Удаление или повреждение соматосенсорных зон коры приводит к нарушению способности локализовать тактильные ощущения, а их электростимуляция вызывает ощущение давления, прикосновения, вибрации и зуда.
|
||
|
|
Последнее изменение этой страницы: 2016-04-18; просмотров: 489; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.33 (0.012 с.) |