

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Метаболизм амидного азота глутамина в почках
Содержание книги
- Структура и функция аминокислот с полярным (заряженным положительно) радикалом. Роль радикалов гистидина в гемоглобине и ферментах. Роль радикалов лизина в структурах коллагена и эластина. Гистоны.
- Методы идентификации и разделения аминокислот
- Химическая модификация аминокислотных радикалов в белках и их роль в построении структуры и функции белков.
- Структура и функции сложных белков: гликопротеиды, протеогликаны, нуклеопротеиды, фосфопротеиды, липопротеиды, хромопротеиды, металлопротеиды.
- Регуляция активности ферментов: неспецифическая, аллостерическая, регуляция на уровне индукции и репрессии, гормональная регуляция.
- Распределение ферментов в клетках, тканях, органах и биологических жидкостях. Мультиферментные комплексы, преимущества организации
- Наследственные энзимопатии - это заболевания, вызванные наследственными нарушениями биосинтеза ферментов или их структуры и функции.
- Структура, классификация и роль углеводов в организме, пищеварение в жкт у новорождённых и детей более старших возрастов, всасывание продуктов гидролиза
- Биологическая роль углеводов
- Аэробный гликолиз: стадия, ферменты и коферменты, биоэнергетика значение для организма. Челночные механизмы транспорта атома водорода в митохондрии и их роль в поддержании аэробного гликолиза.
- Пути вхождения углеводов в гликолиз. Полное окисление фруктозы (начиная с фруктокиназной реакции). Биоэнергетика процесса, фруктозурия.
- Гликогенез и гликогенолиз в печени. Роль печени в поддержании нормального уровня глюкозы в крови.
- Роль печени в поддержании нормального уровня содержания глюкозы в крови.
- Наследственные патологии углеводного обмена: гликогенозы, фруктозурия, галактоземия, непереносимость лактозы и др.
- Липиды организма: классификация, строение, физико-химические свойства, биологическая роль. Пищеварение липидов в жкт новорожденных
- Энтерогепатический цикл холестерола и желчных кислот.
- Синтез гликогена из глицерола в печени.
- Использование кетоновых тел .
- Роль гормонов в регуляции липидного обмена. Патология липидного обмена (жировая инфильтрация печени, атеросклероз, ожирение, наследственные заболевания), причины возникновения и последствия.
- Мембрана эндоплазматического ретикулума (ЭР)
- Переваривание белков в жкт, биосинтез соляной кислоты и пепсиногена в желудке. Протеазы панкреаса и кишечника. Активирование зимогенов в жкт, специфика действия каждой из протеаз.
- Метаболизм аргинина. Окисление аргинина до конечных продуктов, биоэнергетика процесса. Орнитиновый цикл.
- Интеграция обмена аминокислот с обменом других соединений. Синтез глюкозы из глутамата в печени.
- Метаболизм амидного азота глутамина в почках
- Наследственные патологии обмена фенилаланина, тирозина,лейцина,цистеина. Изменения в белковом обмене при сахарном диабете
- Обмен хромопротеидов: синтез и распад гемма. Обтурационная, паренхиматозная и гемолитическая желтуха. Порфирии.
- Обмен нуклеопротеидов: синтез и распад пуринов.
- Этапы реализации генетической информации: репликация, транскрипция, трансляция.
- Особенности метаболизма лейкоцитов.
- Свертывание крови: внешние и внутренние пути.
- Перенос углекислого газа кровью от тканей к легким
- Гипоксия при патологических процессах, нарушающих снабжение или утилизацию кислорода тканями.
- Нарушения кислотно-основного равновесия
- Нарушения общего объема крови
- Патологические изменения эритроцитов
- Распределение в организме и участие в бихимических реакциях мкро и микроэлементов: наприя,калия, кальция,магния,кобольта,железа,цинка,фосфора,хлора и фтора.
- А. Механизм действия гидрофильных гормонов
- А. Механизм действия липофильных гормонов
- Гормоны щитовидной железы,влияние на обмен в-в. Гипо и гипертиреозы. Механизмы возникновения и последствия.
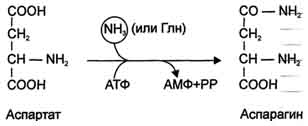
Ещё одной реакцией обезвреживания аммиака в тканях можно считать синтез аспарагина под действием аспарагинсинтетазы.
Наиболее значительные количества аммиака обезвреживаются в печени путём синтеза мочевины. В первой реакции процесса аммиак связывается с диоксидом углерода с образованием карбамоилфосфата, при этом затрачиваются 2 молекулы АТФ. Реакция происходит в митохондриях гепатоцитов под действием фермента карбамоилфос-фатсинтетазы I. Карбамоилфосфатсинтетаза II локализована в цитозоле клеток всех тканей и участвует в синтезе гшримидиновых нуклеотидов (см. раздел 10). Карбамоилфосфат затем включается в орнитиновый цикл и используется для синтеза мочевины.
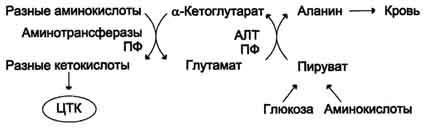
Из мышц и кишечника избыток аммиака выводится преимущественно в виде аланина. Поэтому в мышцах существует ещё один путь выведения азота.
Аминогруппы разных аминокислот посредством реакций трансаминирования переносятся на пируват, основным источником которого служит процесс окисления глюкозы.
Мышцы выделяют особенно много аланина в силу их большой массы
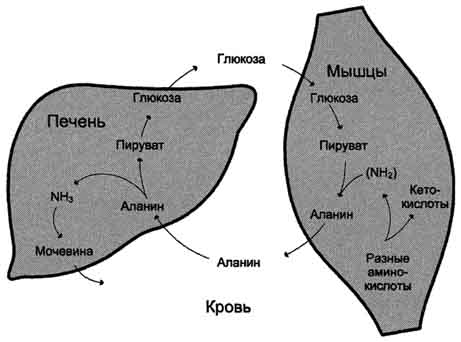
Образовавшийся аланин поступает в печень, где подвергается непрямому дезаминированию. Выделившийся аммиак обезвреживается, а пируват включается в глюконеогенез. Глюкоза из печени поступает в ткани и там, в процессе гликолиза, опять окисляется до пирувата
Образование аланина в мышцах, его перенос в печень и перенос глюкозы, синтезированной в печени, обратно в мышцы составляют глюкозо-аланиновый цикл, работа которого сопряжена с работой глюкозо-лактатного цикла
Глюкозо-аланиновый цикл.
в печени происходит окончательное обезвреживание аммиака – образование мочевины.
Орнитиновый цикл
Основным механизмом обезвреживания аммиака в организме является биосинтезмочевины. Последняя выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного, обмена. Основным и, возможно, единственным местом синтеза мочевины является печень. На первом этапе синтезируется карбамоилфосфат – метаболически активная форма аммиака.
К настоящему времени открыты три разных пути синтеза карбамоилфосфата:
Первую необратимую реакцию катализирует регуляторный фермент – аммиакзависимая карбамоилфосфатсинтетаза:

Реакция требует затраты двух молекулАТФ, открыта в митохондрияхклетокпечени и используется преимущественно для синтеза аргинина и мочевины.
Вторую, также необратимую, реакцию катализирует глутаминзависимая карбамоилфосфатсинтетаза:

Данная реакция открыта в цитозоле клеток животных и требует наличия ионов Mg2+. Следует указать, что благодаря включению гидролитической стадии она используется преимущественно для синтеза пиримидиновых нуклеотидов
Третью обратимую реакцию катализирует карбаматкиназа:

Реакция открыта у разных микроорганизмов и, возможно, используется скорее для ресинтеза АТФ, чем для синтеза карбамоилфосфата.
На втором этапе цикла мочевинообразования происходит конденсация карбамоилфосфата и орнитина с образованием цитруллина; реакцию катализирует орнитин-карбамоилтрансфераза
На следующей стадии цитруллин превращается в аргинин в результате двух последовательно протекающих реакций. Первая из них, энергозависимая,– это конденсация цитруллина и аспарагиновой кислоты с образованием аргининосукцината. Аргининосукцинат распадается в следующей реакции на аргинин и фумарат при участии другого фермента – аргининосукцинатлиазы. На последнем этапе аргинин расщепляется на мочевину и орнитин под действием аргиназы.
Суммарная реакция синтеза мочевины без учета всех промежуточных продуктов может быть представлена в следующем виде:

|




