

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Мотивация и подкрепление - основа голографических построений функций мозга
По аналогии с физической теорией голографии сигнализация о потребности и активируемая ею центральная архитектоника доминирующей мотивации может рассматриваться в качестве опорной волны. Нейроны мозга, вовлеченные в доминирующую мотивацию, в паттерне своей ритмической пачкообразной активности отражают как лежащую в основе доминирующей мотивации потребность, т.е. отклонение результата от уровня, определяющего оптимальную жизнедеятельность, так и свойства потребного подкрепляющего раздражителя - результата поведения.
При подкреплении к этим нейронам мозга устремляются потоки афферентных возбуждений от рецепторов, на которые действуют различные параметры подкрепляющих раздражителей. Подкрепление оказывает специфическое действие на нейроны, вовлеченные в доминирующую мотивацию. Оно переводит их пачкообразную активность в регулярную. С голографических позиций эта афферентации может рассматриваться как предметная волна (Рис. 6).
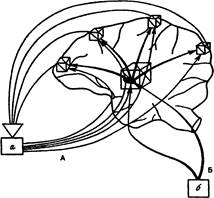
Рис. 6. Голографические аналогии взаимодействия мотивации и подкрепления на структурах мозга: а - подкрепление, б - метаболическая потребность. А - «предметная» сигнализация о подкреплении; Б - «опорная» нервная и гуморальная сигнализация о потребности. Афферентации о метаболической потребности выступает в аналогии с физической «опорной» волной. Афферентации о параметрах результата поведения выступает в форме «предметной» волны.
Афферентация, поступающая в мозг от интерорецепторов и взаимодействующая на структурах мозга с афферентацией от многочисленных экстеро- и интерорецепторов, благодаря распространению возбуждений по перехватам Ранвье нервных волокон, имеет волновую природу. Кроме того, она модулируется "двоичным кодом" результата действия.
Взаимодействие предметной волны с опорной волной создает в структурах мозга своеобразный «отпечаток» - «энграмму подкрепления». Энграмма подкрепления «проявляется» доминирующей мотивацией всякий раз при очередном возникновении соответствующей потребности.
Указанные процессы взаимодействия мотивации и подкрепления генерализованно представлены в различных подкорковых структурах и в коре больших полушарий. Благодаря генерализованному распределению по структурам мозга возбуждений, обусловленных доминирующей мотивацией, экстирпация отдельных участков мозга не нарушает существенно системную организацию поведенческого акта. При наличии сохраненного в гипоталамической области «пейсмекера» доминирующей мотивации, обусловленные им восходящие активирующие возбуждения распространяются по оставшимся интактным структурам мозга, формируя соответствующую мотивацию и поведенческий акт.
Подтверждением такой точки зрения служат наши эксперименты (Судаков К. В., 1990), показавшие, что электролитические разрушения медиальных ядер таламуса и некоторых структур ретикулярной формации ствола мозга меняют высокочастотную низкоамплитудную электрическую активность в коре мозга у голодных животных на медленную, высокоамплитудную. В других же отделах мозга при этом по-прежнему сохраняется обусловленная пищевой потребностью реакция десинхронизации ЭЭГ. Только при двустороннем разрушении «пейсмекера» пищевой мотивации - латеральных отделов гипоталамуса, так же как и после натурального кормления животных, во всех без исключения отделах мозга низкоамплитудная, высокочастотная электрическая активность голодного состояния сменяется медленноволновой, высокоамплитудной. Все это указывает на то, что при наличии активно возбужденного «пейсмекера» в латеральных отделах гипоталамуса даже при разрушении других вовлеченных в эту мотивацию структур мозга, доминирующая пищевая мотивация продолжает проявляться в электрической активности сохраненных структур мозга.
Многочисленные поведенческие эксперименты также указывают на то, что мотивации полностью исчезают только при двустороннем разрушении мотивациогенных отделов гипоталамуса. При разрушениях других отделов лимбических образований мозга и коры больших полушарий при наличии интактного «пейсмекера» в гипоталамической области они сохраняются, хотя внешне могут проявляться в форме сниженной или, наоборот, усиленной поведенческой активности животных. А это значит, что при сохранности доминирующей мотивации у животных может быть не нарушен выработанный ранее условнорефлекторный навык или образован новый.
В публикации (Судаков К. В., 1993) мы показали, что условнорефлекторное возбуждение, наряду с мотивацией и подкреплением, включается в архитектонику аппарата акцептора результата действия, широко распространенную по структурам мозга. Из этого также следует, что удаление значительной части мозга может существенно не сказаться на условнорефлекторной деятельности. К тому же, если иметь в виду сформулированную П. К. Анохиным (1968) конвергентную теорию замыкания условного рефлекса, вопрос становится еще более ясным. Согласно этой теории процесс формирования «временных условнорефлекторных связей» разыгрывается на одних и тех же нейронах мозга и, особенно, коры больших полушарий, на которые распространяются восходящие к ним из подкорковых образований мотивационные, сенсорные и подкрепляющие возбуждения. А это значит, что голографический принцип организации функциональных систем находит отражение в деятельности отдельных нейронов мозга и, возможно, даже в молекулярных процессах, разыгрывающихся в отдельных нейронах мозга. Отсюда можно думать, что при сохранении даже ограниченного числа нейронов мозга при сохраненном «пейсмекере» мотивации условнорефлекторный акт может быть сформирован.
Приведенные материалы позволяют с новых позиций объяснить представления И. П. Павлова о «ядре» и «рассеянных элементах» анализаторов (1949) и представления К. Лешли (Lashley K. S., 1958) о эквипотенциальности коры.
Удаление того или иного участка мозга при сохранности доминирующей мотивации не влияет существенно на проявления условнорефлекторной деятельности. Основные компоненты системной организации поведенческого акта - афферентный синтез, принятие решения, акцептор результата действия и эфферентный синтез - формируются в этом случае за счет оставшихся структур мозга, на которые распространяются мотивационные возбуждения и подкрепление.
Следует однако подчеркнуть, что отмеченные закономерности отражения врожденной доминирующей мотивации в деятельности центральных и периферических структур проявляются отчетливо только в жестких условиях эксперимента: у животных, находящихся под наркозом, или в тех случаях, когда они мягко фиксированы в станке, изолированы от посторонних воздействий и не совершают активной поведенческой деятельности.
Иная картина наблюдается в том случае, когда животные находятся в изменяющихся условиях окружающей их среды или в случае выработки на основе доминирующих биологических мотиваций специальных навыков.
В опытах А. В. Котова (Судаков К. В. и Котов А. В., 1985) характер распределения импульсной активности нейронов у кроликов изучали в различных экспериментальных условиях: в состоянии голода и насыщения и в ситуациях, при которых животные при наличии у них доминирующей мотивации голода были ограничены в достижении полезного результата, т. е. в удовлетворении исходной потребности. Например, пища находилась рядом с животными, но они, будучи иммобилизированными, не имели возможности до нее дотянуться.
Обнаружено, что в условиях создания препятствий на пути удовлетворения пищевой потребности животные начинали проявлять различные типы классической смещенной активности в виде агрессивнооборонительного, комфортного и других видов поведения. Анализ характеров электрической активности нейронов соматосенсорной коры и «центров голода» латерального гипоталамуса у животных в различных функциональных состояниях показал, что в «центрах голода» латерального гипоталамуса после суточной пищевой депривации преобладали клетки с бимодальным и полимодальным распределениями значений межимпульсных интервалов.
При насыщении у кроликов наблюдалось моно- и полимодальное доминирование межимпульсных интервалов - 25-30, 1500-3000 мс и полимодальное - 250-500, 3000-5000, 6000 мс и выше. При агрессивном поведении в латеральном гипоталамусе преобладали нейроны с мономодальным и, особенно, полимодальным распределением межимпульсных интервалов. В сенсомоторной коре при этом нейроны с мономодальным распределением практически не были представлены. Здесь преобладали нейроны с полимодальным распределением межимпульсных интервалов.
При комфортном поведении среди нейронов латерального гипоталамуса преобладали клетки с бимодальным доминированием межимпульсных интервалов. Частично такие же нейроны выявлены и в сенсомоторной коре.
Приведенный пример показывает, что временные распределения паттерна межимпульсной активности нейронов, включенных в доминирующую мотивацию, в процессе активной поведенческой деятельности животных динамически изменяются в зависимости от различных условий достижения ими потребных результатов.
Смена картины распределения межимпульсных интервалов у нейронов мозга, вовлеченных в доминирующую мотивацию, отчетливо наблюдается и на разных стадиях обучения животных, например при выработке у них условных рефлексов. В этом случае в изменении свойств нейронов, включенных в доминирующую мотивацию, первостепенную роль играет подкрепление (Журавлев Б. В. и Умрюхин Е. А., 1993).
Все это указывает на то, что голографические свойства врожденной биологической мотивации существенно изменяются в зависимости от изменения состояния животного, его реакций на разнообразные внешние раздражители, успешности удовлетворения ведущей потребности, наконец в зависимости от опыта, обучения.
Под влиянием постоянно возбужденного соответствующей потребностью гипоталамического «пейсмекера» генерализованные потоки восходящих возбуждений широко распространяются по значительной массе мозга. За счет реверберации корково-подкорковых возбуждений наблюдается расчленение потоков импульсной активности нейронов различного уровня мозга, вовлеченных в доминирующую мотивацию, усиливающие частотно-модулирующий характер опорной волны. Эти реверберирующие влияния охватывают не только структуры мозга, но и периферические рецепторы. В результате - голографический экран доминирующей мотивации, на который проецируются афферентные возбуждения, связанные с подкреплением, включает как соответствующий рецепторный аппарат, так и структуры мозга.
Мотивационное возбуждение производит избирательную настройку нейронов различного уровня мозга и периферических рецепторов к подкрепляющим воздействиям. В результате формируется своеобразный информационный «фильтр» для поступающих в мозг возбуждений, вызванных действием подкрепляющих факторов. Возбуждения, поступающие в мозг от различных параметров подкрепления, адресуются к различным нейронам, находящимся на разных уровнях мозга, и при каждом воздействии оставляют след в их деятельности. Благодаря этому в любой момент времени входное воздействие, связанное с возбуждением периферических рецепторов, сопоставляется с конфигурацией возбуждения, существовавшего до его прихода в различных структурах мозга.
Установлено, что подкрепление изменяет свойства нейронов, ранее включенных в исходную доминирующую мотивацию. Изменяются их конвергентные свойства и чувствительность к нейромедиаторам и нейропептидам (Судаков К. В., 1996). Подкрепление изменяет молекулярную структуру нейронов мозга. После предварительного подкрепления, как будет показано ниже, нейроны разных отделов мозга начинают экспрессировать специальные белковые молекулы. Отсюда можно думать, что подкрепление изменяет и оптические свойства генетического материала ДНК. Можно предполагать, что жидкие кристаллы ДНК играют роль своеобразной оптической линзы в мозге. Кристаллы ДНК обладают четко выраженными оптическими свойствами (Франк-Каменецкий М. Д., 1988). Аминокислоты (за исключением глицина) так же оптически активны, т.е. в водных или подкисленных растворах вращают плоскость поляризованного света вправо или влево в зависимости от природы аминокислоты.
Преломляющую функцию в мозге могут играть структурно организованные коллоиды межклеточного пространства и глии - мицеллы.
К. Прибрам (1975) в происхождении голограммы мозга ведущее значение придает не нервным импульсациям аксонов, а микроструктуре медленных потенциалов, которые формируются постсинаптическими и дендритными процессами.
Как видно, вопросы организации голографического экрана мозга все еще гипотетичны и требуют дальнейшего изучения.
|




