Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Функциональная морфология нейрона
Нейрон состоит из клеточного тела (перикариона) и отростков, обеспечивающих проведение нервных импульсов - дендритов, приносящих импульсы к телу нейрона, и аксона (нейрита), несущего импульсы от тела нейрона (рис. 52; рис. 53). Тело нейрона (перикарион) включает ядро и окружающую его цитоплазму (за исключением входящей в состав отростков). Перикарион содержит синтетический аппарат нейрона, а его плазмолемма осуществляет рецепторные функции, так как на ней находятся многочисленные нервные окончания (синапсы), несущие возбуждающие и тормозные сигналы от других нейронов. Ядро нейрона – обычно одно, крупное, округлое, светлое, с мелкодисперсным хроматином (преобладанием эухроматина), одним, иногда 2-3 крупными ядрышками. Эти особенности отражают высокую активность процессов транскрипции в ядре нейрона. Около ядрышка в нейронах у лиц женского пола часто выявляется тельце Барра – крупная глыбка хроматина, содержащая конденсированную Х-хромосому (особенно заметна в клетках коры полушарий большого мозга и симпатических нервных узлов). Цитоплазма нейрона богата органеллами и окружена плазмолеммой, которая обладает способностью к проведению нервного импульса (распространению деполяризации) вследствие локального тока Na+ в цитоплазму и К+ из нее через потенциал-зависимые мембранные ионные каналы. Плазмолемма содержит Na+-K+ насосы, которые поддерживают необходимые градиенты ионов.
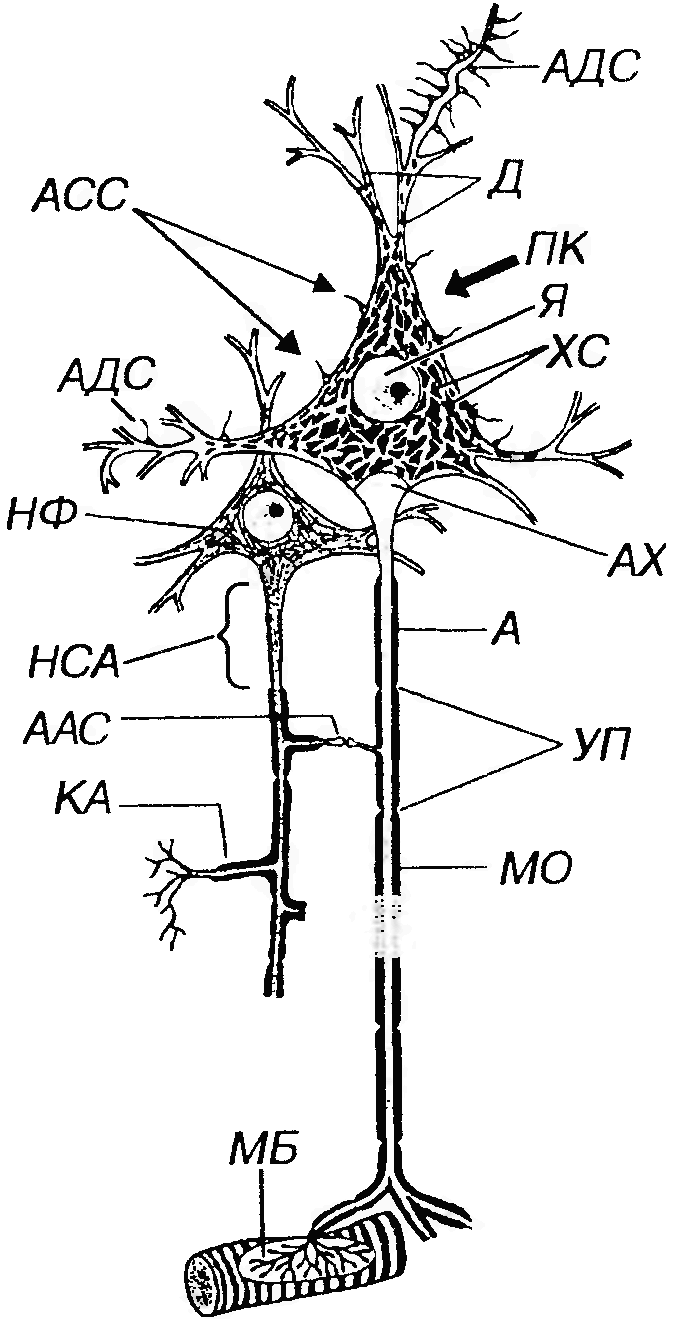
Рис. 52. Строение мультиполярного нейрона (по Rohen J. W., Lutjen - Drecoll E., 1982). ПК - перикарион, Я - ядро с ядрышком, ХС - хроматофильная субстанция, НФ - нейрофибриллы (агрегаты элементов цитосклета), Д – дендриты, А - аксон, НСА - начальный сегмент аксона, АХ - аксонный холмик, КА - коллатерали аксона, МО - миелиновая оболочка, УП - узловые перехваты, МБ - моторная бляшка (двигательное нервное окончание на волокне поперечнополосатой мышцы), Синапсы (С): АДС - аксо-дендритический, АСС - аксо-соматический, ААС - аксо-аксональный.
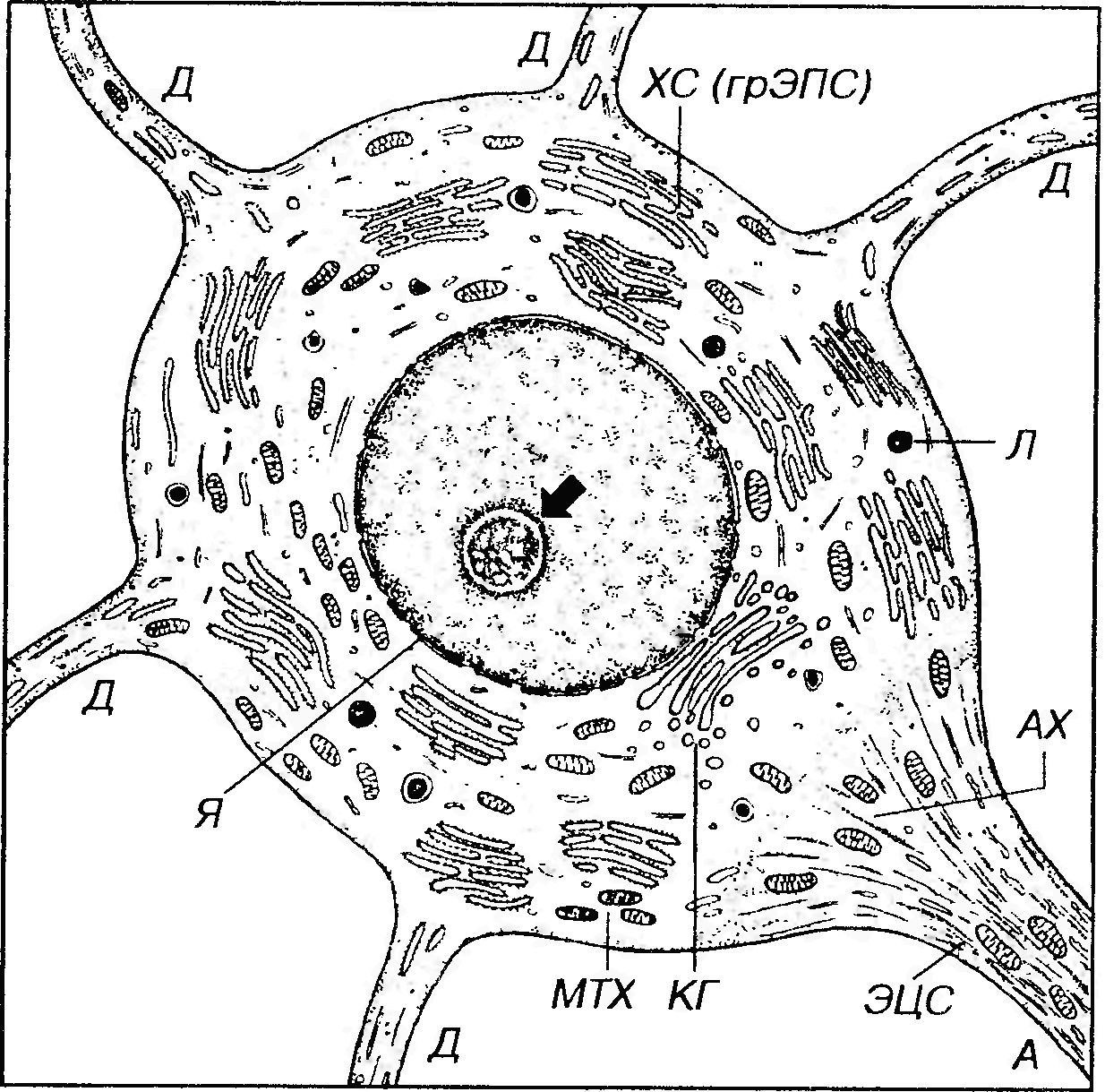
Рис. 53. Ультраструктурная организация нейрона. Я - ядро (ядрышко показано стрелкой), ХС - хроматофильная субстанция, ЭЦС - элементы цитоскелета (нейротрубочки, нейрофиламенты), МТХ - митохондрии, КГ - комплекс Гольджи, Л - лизосомы, Д - дендриты, А - аксон, АХ - аксонный холмик.
грЭПС хорошо развита, ее цистерны часто образуют отдельные комплексы из параллельно лежащих уплощенных анастомозируюших элементов, которые на светооптическом уровне при окраске анилиновыми красителями имеют вид базофильных глыбок, в совокупности получивших название хроматофильной субстанции (вещества, или телец Ниссля, тигроидного вещества, тигроида). Характер распределения и размеры комплексов цистерн грЭПС (хроматофильной субстанции) варьируют в отдельных типах нейронов (наиболее крупные обнаруживаются в мотонейронах) и зависят от их функционального состояния. При длительном раздражении или повреждении нейрона комплексы цистерн грЭПС распадаются на отдельные элементы, что на светооптическом уровне проявляется исчезновением телец Ниссля (хроматолиз, тигролиз). аЭПС образована трехмерной сетью анастомозирующих цистерн и трубочек, участвующих в синтетических процессах и внутриклеточном транспорте веществ. Комплекс Гольджи хорошо развит и состоит из множественных диктиосом, расположенных обычно вокруг ядра. Митохондрии – очень многочисленны и обеспечивают высокие энергетические потребности нейрона, связанные со значительной активностью синтетических процессов, проведением нервных импульсов, деятельностью ионных насосов. Они обычно имеют палочковидную форму и характеризуются быстрым изнашиванием и обновлением (коротким жизненным циклом). Лизосомальный аппарат (аппарат внутриклеточного переваривания) обладает высокой активностью и представлен эндосомами и многочисленными лизосомами различных размеров. Интенсивные процессы аутофагии обеспечивают постоянное обновление компонентов цитоплазмы нейрона. При дефектах некоторых лизосомальных ферментов в цитоплазме нейронов накапливаются непереваренные продукты, что нарушает их функции и вызывает болезни накопления, например, ганглиозидоз (болезнь Тэй-Закса). Включения в цитоплазме нейрона представлены липидными каплями, гранулами липофусцина, которыйявляется даже в нейронах плодов и (нейро)меланина. Цитоскелет нейронов хорошо развит и представлен всеми элементами – микротрубочками (нейротрубочками), микрофиламентами и промежуточными филаментами (нейрофиламентами). Они образуют трехмерную опорно-сократительную сеть, играющую важную роль в поддержании формы этих клеток и, в особенности, их длинного отростка – аксона (нейрита). Многочисленные промежуточные филаменты (нейрофиламенты) связаны друг с другом и с нейротрубочками поперечными мостиками; при фиксации они склеиваются в пучки, которые окрашиваются солями серебра. Такие образования (фактически являющиеся артефактами) на светооптическом уровне описаны под названием нейрофибрилл – нитей толщиной 0.5-3 мкм, образующих сеть в перикарионе. Микротрубочки (нейротрубочки) и микрофиламенты имеют такое же строение, как и в других клетках. Клеточный центр присутствует во всех нейронах, его главная функция – сборка микротрубочек.
Дендриты проводят импульсы к телу нейрона, получая сигналы от других нейронов через многочисленные межнейронные контакты (аксо-дендритические синапсы), расположенные на них в области особых цитоплазматических выпячиваний – дендритных шипиков. Во многих шипиках имеется особый шипиковый аппарат, состоящий из 3-4 уплощенных цистерн, разделенных участками плотного вещества. Шипики представляют собой лабильные структуры, которые разрушаются и образуются вновь; их число резко падает при старении, а также при снижении функциональной активности нейронов. В большинстве случаев дендриты многочисленны, имеют относительно небольшую длину и сильно ветвятся вблизи тела нейрона. Крупные стволовые дендриты содержат все виды органелл, по мере снижения их диаметра в них исчезают элементы комплекса Гольджи, а цистерны грЭПС сохраняются. Нейротрубочки и нейрофиламенты многочисленны и располагаются параллельными пучками; они обеспечивают дендритный транспорт (рис. 54), который осуществляется из тела клетки вдоль дендритов со скоростью около 3 мм/ч.
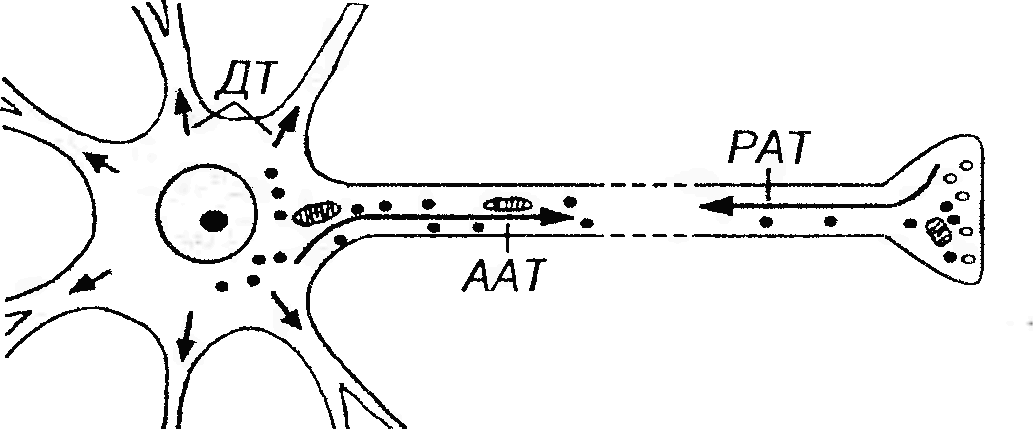
Рис. 54. Процессы транспорта в нейроне. ААТ - антероградный аксонный транспорт (из тела нейрона по аксону) подразделяется на медленный (скорость -1-5 мм/сут.) и быстрый (100-500 мм/сут.). PAT - ретроградный аксонный транспорт (из аксона в тело нейрона) осуществляется со скоростью 100-200 мм/сут. ДТ - дендритный транспорт (из тела клетки по дендритам) происходит со скоростью около 70 мм/сут. Аксон – длинный (у человека от 1 мм до 1,5 м) отросток, по которому нервные импульсы передаются на другие нейроны или клетки рабочих органов (мышц, желез). В крупных нейронах аксон может содержать до 99% объема цитоплазмы. Аксон отходит от утолщенного участка тела нейрона, не содержащего хроматофильной субстанции, – аксонного холмика, в котором генерируются нервные импульсы; почти на всем протяжении он покрыт глиальной оболочкой. Центральная часть цитоплазмы аксона (аксоплазмы) содержит пучки нейрофиламентов, ориентированных вдоль его длины, ближе к периферии располагаются пучки микротрубочек, цистерны аЭПС, элементы комплекса Гольджи, митохондрии, мембранные пузырьки, сложная сеть микрофиламентов. Тельца Ниссля в аксоне отсутствуют. Аксон может по своему ходу давать ответвления (коллатерали), которые обычно отходят от нею под прямым углом. В конечном участке аксон нередко распадается на тонкие веточки (телодендрии). Аксон заканчивается специализированными терминалями (нервными окончаниями) на других нейронах или клетках рабочих органов.
Аксонный транспорт (ток) – перемещение по аксону различных веществ и органелл (рис. 54); разделяется на антероградный (прямой – из тела нейрона по аксону) и ретроградный (обратный – из аксона в тело нейрона). Вещества переносятся в цистернах аЭПС и пузырьках, которые перемещаются вдоль аксона благодаря взаимодействию с элементами цитоскелета (главным образом, с микротрубочками посредством связанных с ними сократимых белков – кинезина и динеина); процесс транспорта является Са2+-зависимым. Антероградный аксонный транспорт включает медленный (скорость – 1-5 мм/сут.), обеспечивающий ток аксоплазмы (переносящий ферменты и элементы цитоскелета), и быстрый (100-500 мм/сут.), осуществляющий перенос различных веществ, цистерн грЭПС, митохондрий, пузырьков, содержащих нейромедиаторы. Ретроградный аксонный транспорт (100-200 мм/сут.) способствует удалению веществ из области терминалей, возвращению пузырьков, митохондрий. Предполагается, что за счет аксонного транспорта проникшие в нейрон нейротропные вирусы (герпеса, бешенства, полиомиелита) могут распространяться по нейронным цепям. Феномен транспорта используется для изучения межнейронных связей путем введения маркера в область расположения терминалей или клеточных тел и выявления областей его последующего распространения описанными механизмами. КЛАССИФИКАЦИЯ НЕЙРОНОВ Классификация нейронов осуществляется по трем признакам: морфологическим, функциональным и биохимическим. Морфологическая классификация нейронов учитывает количество их отростков и подразделяет все нейроны на три типа (рис. 55): униполярные, биполярные и мультиполярные. Униполярные нейроны имеют один отросток. По мнению большинства исследователей, в нервной системе человека и других млекопитающих они не встречаются. Некоторые авторы к таким клеткам все же относят амакринные нейроны сетчатки глаза и межклубочковые нейроны обонятельной луковицы.
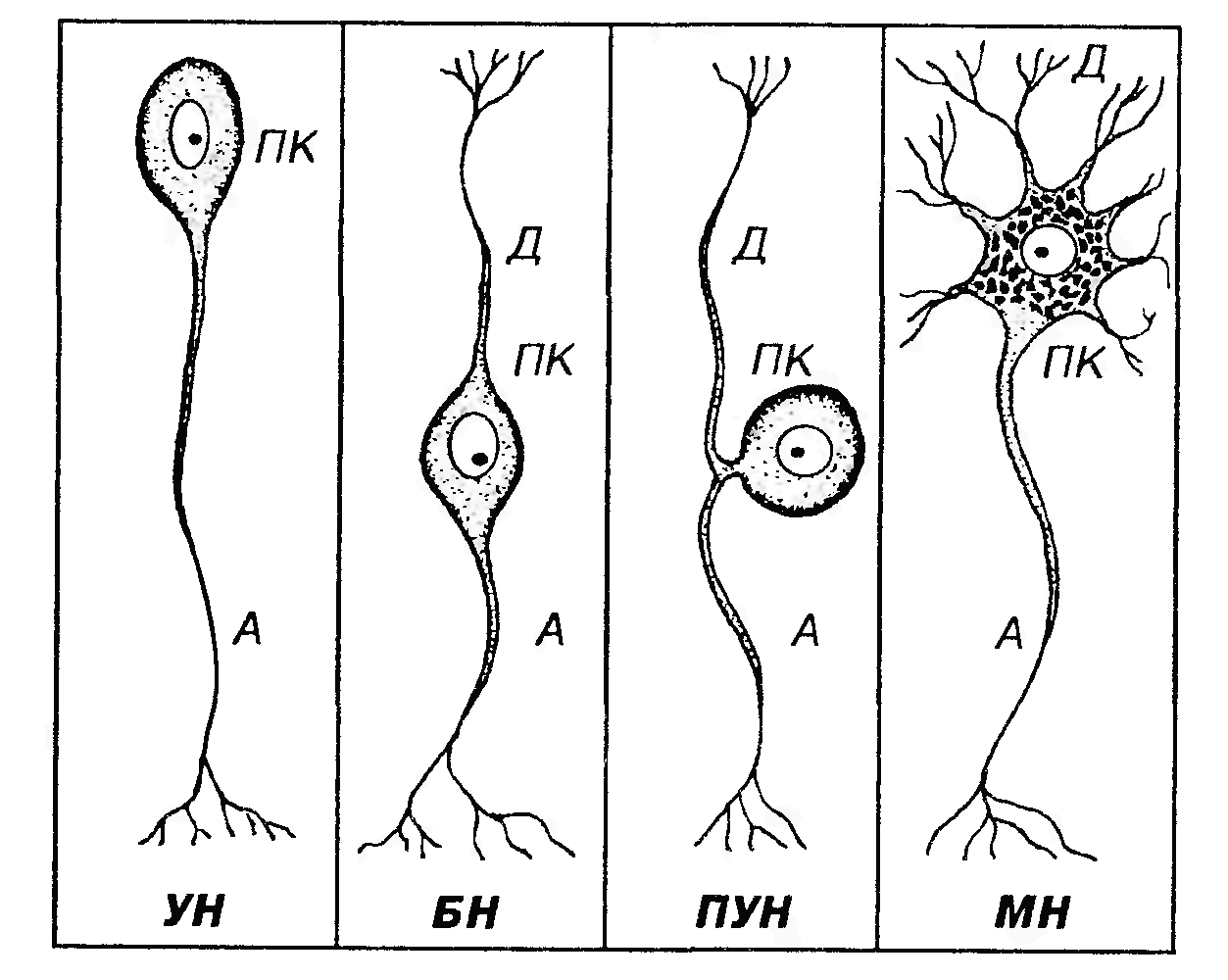
Рис. 55. Морфологическая классификация нейронов. УН - униполярный нейрон, БН - биполярный нейрон, ПУН - псевдоуниполярный нейрон, МН - мультиполярный нейрон, ПК - перикарион, А – аксон, Д - дендрит(ы).
Биполярные нейроны имеют два отростка – аксон и дендрит, обычно отходящие от противоположных полюсов клетки. В нервной системе человека встречаются редко. К ним относят биполярные клетки сетчатки глаза, спирального и вестибулярного ганглиев.
Псевдоуниполярные нейроны – разновидность биполярных, в них оба клеточных отростка (аксон и дендрит) отходят от тела клетки в виде единого выроста, который далее Т-образно делится. Эта клетки встречаются в спинальных и краниальных ганглиях. Мультиполярные нейроны имеют три или большее число отростков: аксон и несколько дендритов. Они наиболее распространены в нервной системе человека. Описано до 80 вариантов этих клеток: веретенообразные, звездчатые, грушевидные, пирамидные, корзинчатые и др. По длине аксона выделяют клетки Гольджи I типа (с длинным аксоном) и клетки Гольджи II типа (с коротким аксоном). Функциональная классификация нейронов разделяет их по характеру выполняемой ими функции (в соответствии с их местом в рефлекторной дуге) на три типа: чувствительные, двигательные и ассоциативные. Чувствительные (афферентные) нейроны генерируют нервные импульсы под влиянием изменений внешней или внутренней среды. Двигательные (эфферентные) нейроны передают сигналы на рабочие органы (скелетные мышцы, железы, кровеносные сосуды). Ассоциативные (вставочные) нейроны (интернейроны) осуществляют связи между нейронами и количественно преобладают над нейронами других типов, составляя в нервной системе около 99,98% от общего числа этих клеток. Биохимическая классификация нейронов основана на химических особенностях нейромедиаторое, используемых нейронами в синаптической передаче нервных импульсов. Выделяют много различных групп нейронов, в частности, холинергические (медиатор – ацетилхолин), адренергические (медиатор – норадреналин), серотонинергические (медиатор – серотонин), дофаминергические (медиатор – дофамин), ГАМК-ергические (медиатор – гамма-аминомасляная кислота, ГАМК), пуринергические (медиатор – АТФ и его производные), пептидергические (медиаторы – субстанция Р, энкефалины, эндорфины, вазоактивный интестинальный пептид, холецистокинин, нейротензин, бомбезин и другие нейропептиды). В некоторых нейронах терминали содержат одновременно два тина нейромедиатора. Распределение нейронов, использующих различные медиаторы, в нервной системе неравномерно. Нарушение выработки некоторых медиаторов в отдельных структурах мозга связывают с патогенезом ряда нервно-психических заболеваний. Так, содержание дофамина снижено при паркинсонизме и повышено при шизофрении, снижение уровней норадреналина и серотонина типично для депрессивных состояний, а их повышение – для маниакальных. МЕЖНЕЙРОНАЛЬНЫЕ КОНТАКТЫ – СИНАПСЫ – это специфические контакты нейронов, обеспечивающие передачу возбуждения от одной нервной клетки к другой. В зависимости от способов передачи возбуждения (так называемого потенциала действия) выделяют химические и электрические синапсы.
Эволюционно более древними и примитивными являются электрические синаптические контакты. Они по строению близки к щелевидным контактам (нексусам) и представляют собой ограниченные области контактов двух соседних нейронов, где расстояние между соседними мембранами сужено до 3-4 нм. В зоне этих сужений имеются белковые мембранные комплексы, аналогичные коннексонам, формирующим пору. Через пору могут свободно диффундировать вода, ионы, низкомолекулярные органические вещества (в том числе и гормоноподобные). Считается, что обмен происходит в обе стороны, но имеются случаи, когда возбуждение передается в одном направлении. Таким образом, возбуждение в таком синапсе в большинстве случаев идет в обе стороны и носит равновероятностный, диффузный (ирритативный) характер. Такие контакты часто встречаются у низших беспозвоночных и хордовых. У млекопитающих электрические контакты имеют большое значение в процессе межнейронных взаимодействий в эмбриональном периоде развития. С дифференцировкой нервной ткани их число убывает. Химические синапсы для передачи возбуждения от одной нервной клетки к другой используют специальные вещества – медиаторы (нейромедиаторы). Кроме медиаторов ими используются и модуляторы. Модуляторы – это специальные химические вещества, которые сами возбуждения не вызывают, но могут либо усиливать, либо ослаблять чувствительность к медиаторам (т. е. модулировать пороговую чувствительность клетки к возбуждению). Каждый нейрон содержит один специфичный для него медиатор и один или несколько модуляторов. Эти вещества накапливаются в синаптических пузырьках, которые отделены от матрикса цитоплазмы мембраной. Форма, размеры, строение пузырьков зависят от содержащегося в них вещества. Любой из химических синапсов обеспечивает однонаправленную передачу возбуждения. В химическом синапсе видны: пресинаптическая зона (пресинаптическое расширение), в основном представляющая собой терминаль аксона (рис. 56); синаптическая щель диаметром 20-50 нм; постсинаптическая зона. Пресинаптическая зона обязательно содержит синаптические пузырьки, элементы цитоскелета (нейротубулы и нейрофиламенты), митохондрии. Часто мембрана, обеспечивающая передачу импульса, имеет хорошо выраженное электронно-плотное вещество. Оно представляет собой скопление мембранных белков, одной из функций которых является направленное выделение содержимого синаптических пузырьков в синаптическую щель. Таким образом, выделение медиаторов осуществляется лишь через строго определенные участки пресинаптической мембраны. Синаптическая щель изолируется от окружающего межклеточного вещества нейроглией (в центральной нервной системе отростками астроцитов). Постсинаптическая зона включает в себя электронно-плотное вещество постсинаптической мембраны, митохондрии. Электронно-плотное вещество также составлено комплексом мембранных белков, в том числе рецепторных (на гликокаликсе), белков ионных каналов и ферментов, расщепляющих медиаторы.
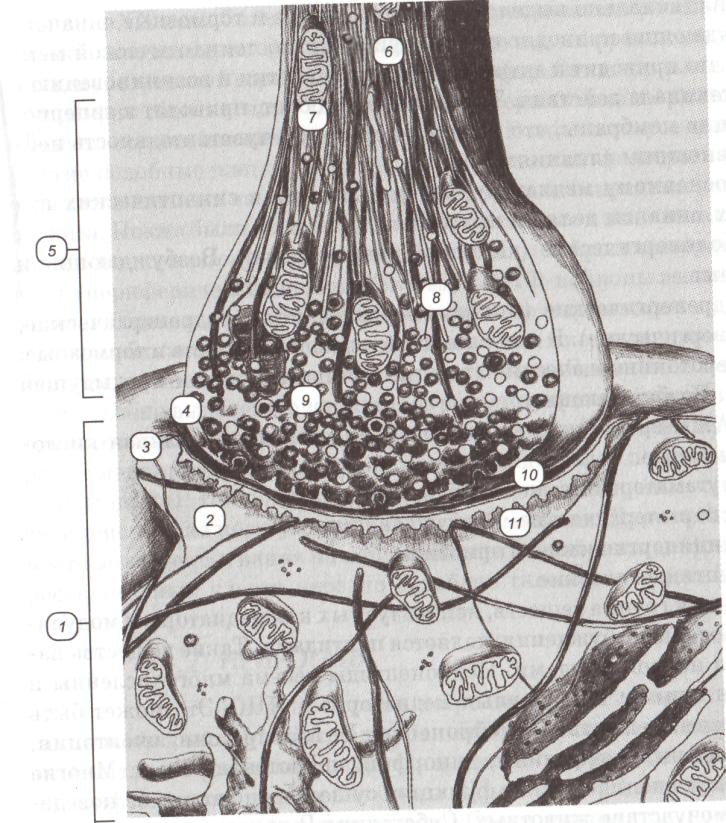
Рис. 56. Принципиальное строение простого химического синапса: 1 - постсинаптическая зона; 2 - постсинаптическая мембрана; 3 - синаптическая щель; 4 - пресинаптическая мембрана; 5 - пресинаптическое расширение; 6 - нейротубулы; 7 - митохондрия 8 - синаптические пузырьки с медиатором; 9 - синаптические с модулятором; 10 - электронно-плотное вещество пресинаптической мембраны; 11 - электронно-плотное вещество постсинаптической мембраны.
В зависимости от того, какие структуры взаимодействуют в синапсе можно выделить следующие виды синапсов: аксо-дендритические (пресинаптическая структура – аксон, постсинаптическая – дендрит); а аксо-аксональные (аксон с аксоном); аксо-соматические (аксон с телом нервной клетки). Функционально выделяют возбуждающие и тормозные синапсы. Возбуждающие приводят к деполяризации постсинаптической мембраны, что приводит к активации нервной клетки и возникновению в ней потенциала действия. Тормозные, наоборот, приводят к гиперполяризации мембраны, что снижает пороговую чувствительность нейрона к внешним влияниям. По основному медиатору, содержащемуся в синаптических пузырьках, синапсы делятся на группы (см. биохимическую классификацию нейронов). В последние годы значительный интерес проявляется к роли оксида азота как к медиатору. Оксид азота модулирует возбуждение в нервных клетках, но само вещество может диффундировать в соседние клетки и влиять на них и без участия синапсов (внесинаптическая нервная передача). В частности, значительная группа нейронов – водителей ритма (пейсмекеров) – имеет NO-активность. Секреторные (эндокринные) нейроциты. Впоследние десятилетия все больше внимания уделяется вопросам нейросекреции. Еще в начале XX века были обнаружены классические нейросекреторные клетки гипоталамуса. Это нейроны, выделяющие в кровь окситоцин и антидиуретический гормон через нейрогипофиз. Они направляют в нейрогипофиз свои аксоны, где формируют терминальные расширения вблизи сосудов (аксовазальные синапсы). В гипоталамусе имеются и другие подобные клетки, но выделяющие релизинг-факторы в срединном возвышении в непосредственной близости от основания ножки гипофиза. Позже были выявлены клетки, способные к выделению гормонов в спинно-мозговую жидкость. Обнаружены нейросекреторные клетки и в периферической нервной системе (МИФ-нейроны вегетативных узлов). В целом нейросекреторные клетки по морфологической организации близки к типичным нейронам. Они имеют сходную с ними систему органелл и синаптических контактов. Выделение нейросекрета подобно выделению медиаторов регулируется возбуждением нейрона. Отличия заключаются в том, что нейросекрет они выделяют наряду с медиатором. Нейросекрет обычно представлен пептидами (нейропептидами). Нейропептиды могут выводиться в кровь и оказывать свое действие на периферические органы-мишени. Другие оказывают местные эффекты, влияя на нервные клетки и нейроглию. Вокруг нейросекреторных клеток нередко ослаблен гистогематический барьер. НЕЙРОГЛИЯ ОБЩИЕ СВЕДЕНИЯ Нейроглия – обширная гетерогенная группа элементов нервной ткани, обеспечивающая деятельность нейронов и выполняющая опорную, трофическую, разграничительную, барьерную, секреторную и защитную функции. В мозге человека содержание глиальных клеток (глиоцитов) в 5-10 раз превышает число нейронов, причем они занимают около половины его объема. В отличие от нейронов, глиоциты взрослого способны к делению. В поврежденных участках мозга они размножаются, заполняя дефекты и образуя глиальные рубцы (глиоз); опухоли из клеток глии (глиомы) составляют 50% внутричерепных новообразований.
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-02-07; просмотров: 248; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.111.125 (0.027 с.) |