Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Функциональная морфология скелетной мышечной ткани
Скелетная (соматическая) мышечная ткань образована пучками поперечнополосатых мышечных волокон, являющихся ее структурно-функциональными единицами. Всего в скелетных мышцах человека содержится порядка 300 млн. мышечных волокон.
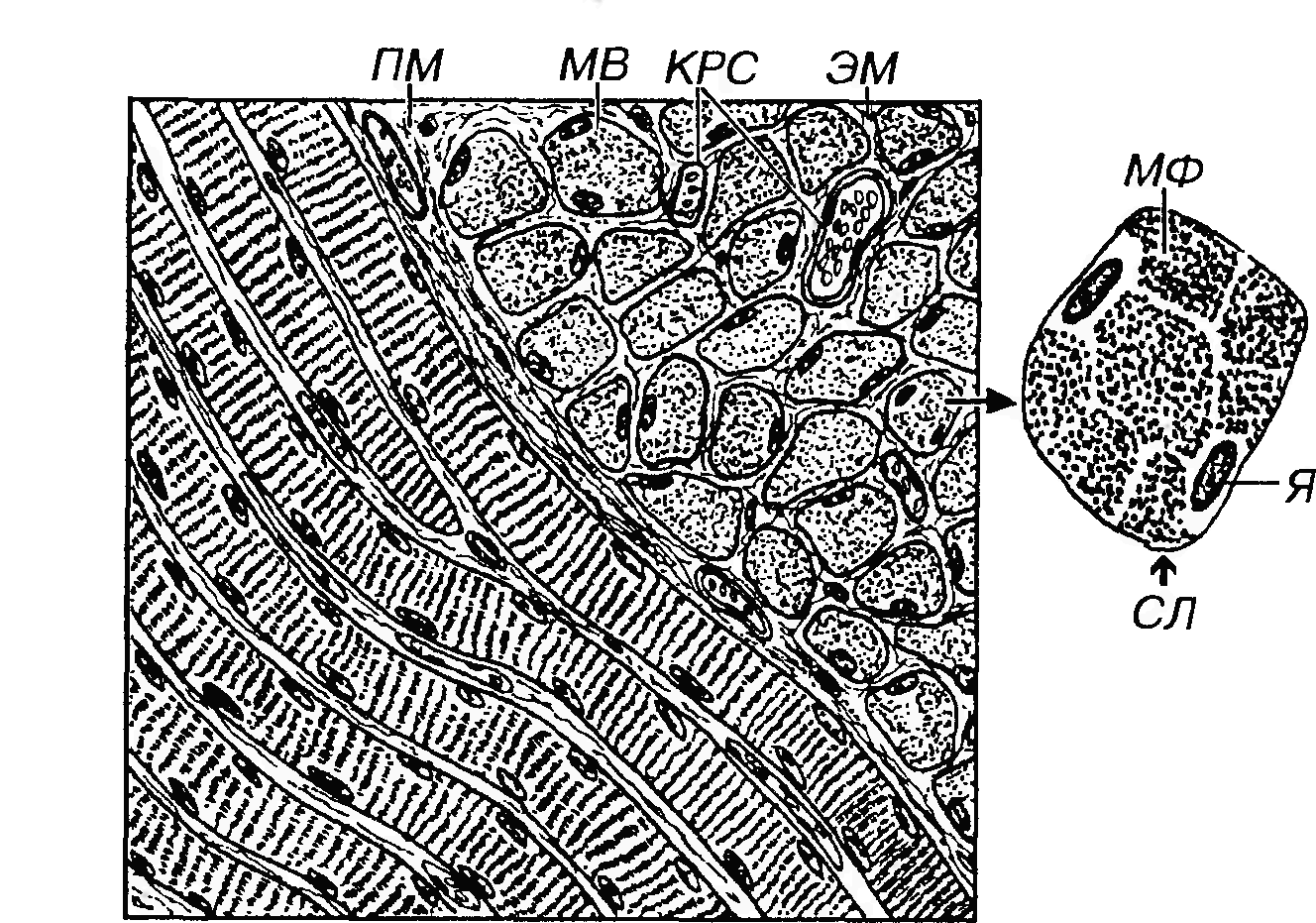
Мышечное волокно скелетной (соматической) мышечной ткани представляет собой цилиндрическое образование диаметром 10-100 мкм (в среднем - 50 мкм) вариабельной длины (до 10-30 см). Мышечные волокна в мышцах образуют пучки, в которых они лежат параллельно и, деформируя друг друга, часто приобретают неправильную многогранную форму (рис. 40).
Рис. 40. Скелетная (соматическая) мышечная ткань. Продольные (слева) и поперечные (справа) разрезы мышечных волокон (MB), между которыми располагается эндомизий (ЭМ). Пучки MB покрыты более толстой соединительнотканной оболочкой - перимизием (ПМ). Кровеносные сосуды (КРС) из ПМ проникают в ЭМ. На поперечном разрезе отдельного мышечного волокна видны сарколемма (СЛ), периферически лежащие ядра (Я) миосимпласта и центрально расположенные миофибриллы (МФ), собранные в группы (поля Конгейма). Диаметр волокон обусловливается: (1) их принадлежностью к определенной мышце (они тонкие в глазных мышцах, толстые в мышцах спины и конечностей), (2) полом (толще у мужчин), (3) возрастом (увеличивается более, чем 10-кратно после рождения), (4) состоянием питания (истончаются при его недостаточности), (5) степенью функциональной нагрузки - волокна утолщаются (гипертрофируются) при усиленной нагрузке и истончаются (атрофируются) при ее снижении и, в особенности, при денервации.
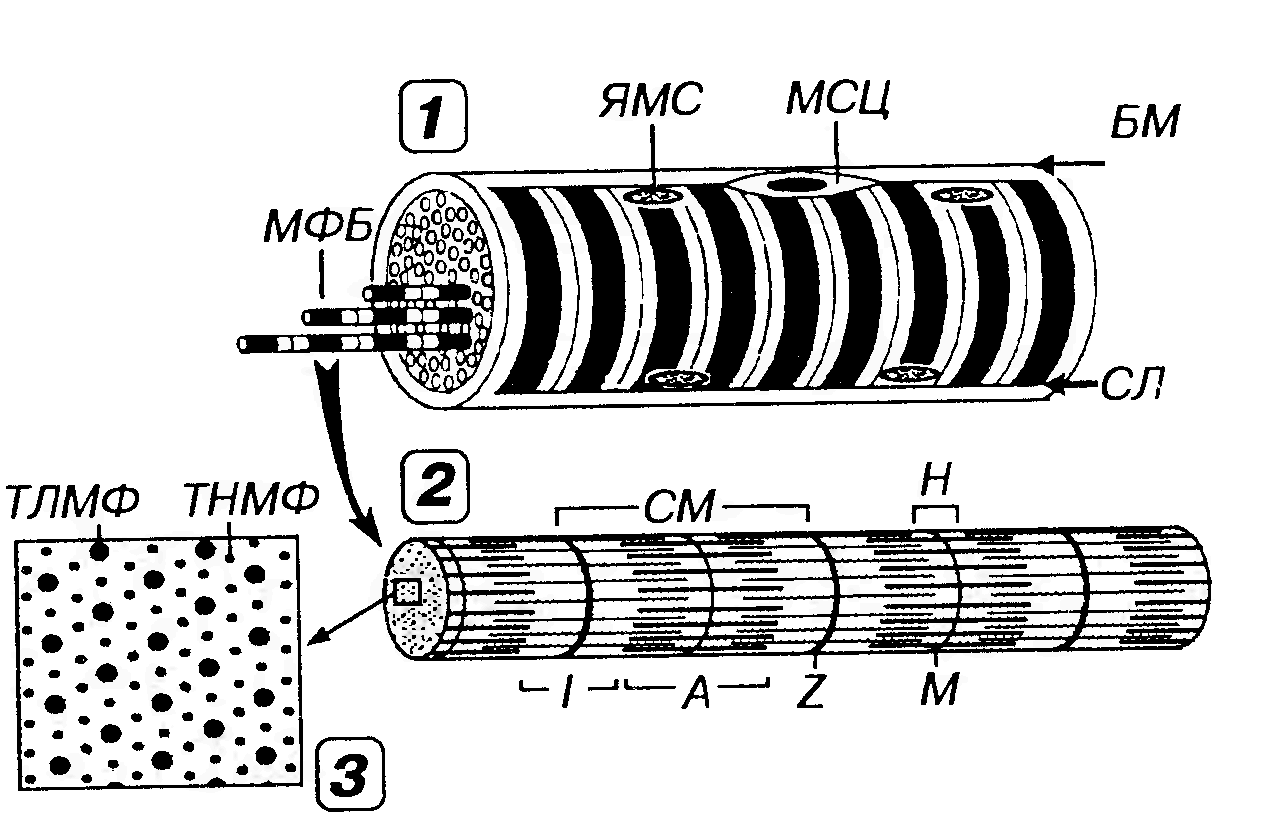
Поперечная исчерченность скелетных мышечных волокон обусловлена чередованием темных А-дисков (анизотропных, обладающих двойным лучепреломлением в поляризованном свете) и светлых 1-дисков (изотропных, не обладающих двойным лучепреломлением). Каждый диск I рассекается надвое тонкой темной Z -линией (от немецкого Zwischenscheibe - промежуточный диск), называемой также телофрагмой. В середине A-диска определяется светлая зона - полоска Н (от немецкого helle - светлый), через центр которой проходит М-линия - мезофрагма (рис. 41).
Рис. 41. Строение скелетного мышечного волокна (1), миофибриллы (2) и расположение в последней миофиламентов (3). В мышечном волокне (MB) в целом и каждой миофибрилле (МФ), входящей в его состав, выявляются чередующиеся темные анизотропные А-диски (А) и светлые изотропные 1-диски (I). Последние рассекаются надвое телофрагмой, или Z -линией (Z), а в середине первых определяется светлая полоска Н (Н), через центр которой проходит М-линия (М). СМ - саркомер, ЯМС - ядра миосимпласта, МСЦ - миосателлитоцит, БМ - базальная мембрана, СЛ - сарколемма. В пределах СМ каждый толстый миофиламент (ТЛМФ) окружен шестью тонкими миофиламентами (ТНМФ).
Компонентами мышечного волокна являются: (1) миосимпластическая часть (которая занимает основной его объем и ограничена сарколеммой) и (2) миосателлитоциты - мелкие уплощенные клетки, прилежащие к поверхности миосимпласта и располагающиеся в углублениях его сарколеммы. Снаружи сарколемма покрыта толстой базальной мембраной, в которую вплетаются ретикулярные волокна. Некоторые авторы сарколеммой волокна скелетной мышечной ткани именуют не его плазмолемму, а совокупность плазмолеммы и базальной мембраны, что является отражением представлений прежних лет (до изобретения электронного микроскопа), когда эти две отдельные структуры воспринимали на светооптическом уровне, как единое образование. Миосимпластическая часть мышечного волокна включает от нескольких сотен до нескольких тысяч ядер, лежащих на периферии под сарколеммой, и саркоплазму, образующую его центральную часть. Ядра миосимпласта - сравнительно светлые, с 1-2 ядрышками, диплоидные, овальные, уплощенные, длиной 10-20 мкм. Ориентированы длинной осью вдоль волокна и располагаются на расстоянии около 5 мкм друг от друга. При резком сокращении волокон они могут укорачиваться, деформироваться и штопорообразно скручиваться. Содержание ядер несколько выше в красных волокнах по сравнению с белыми. Саркоплазма миосимпласта содержит все органеллы общего значения (за исключением центриолей) и некоторые специальные органеллы, а также включения. Эти структуры образуют несколько функциональных аппаратов: 1) сократительный, 2) передачи возбуждения (с сарколеммы на сократительный аппарат), 3) опорный, 4) энергетический, 5) синтетический, б) лизосомальный (аппарат внутриклеточного переваривания).
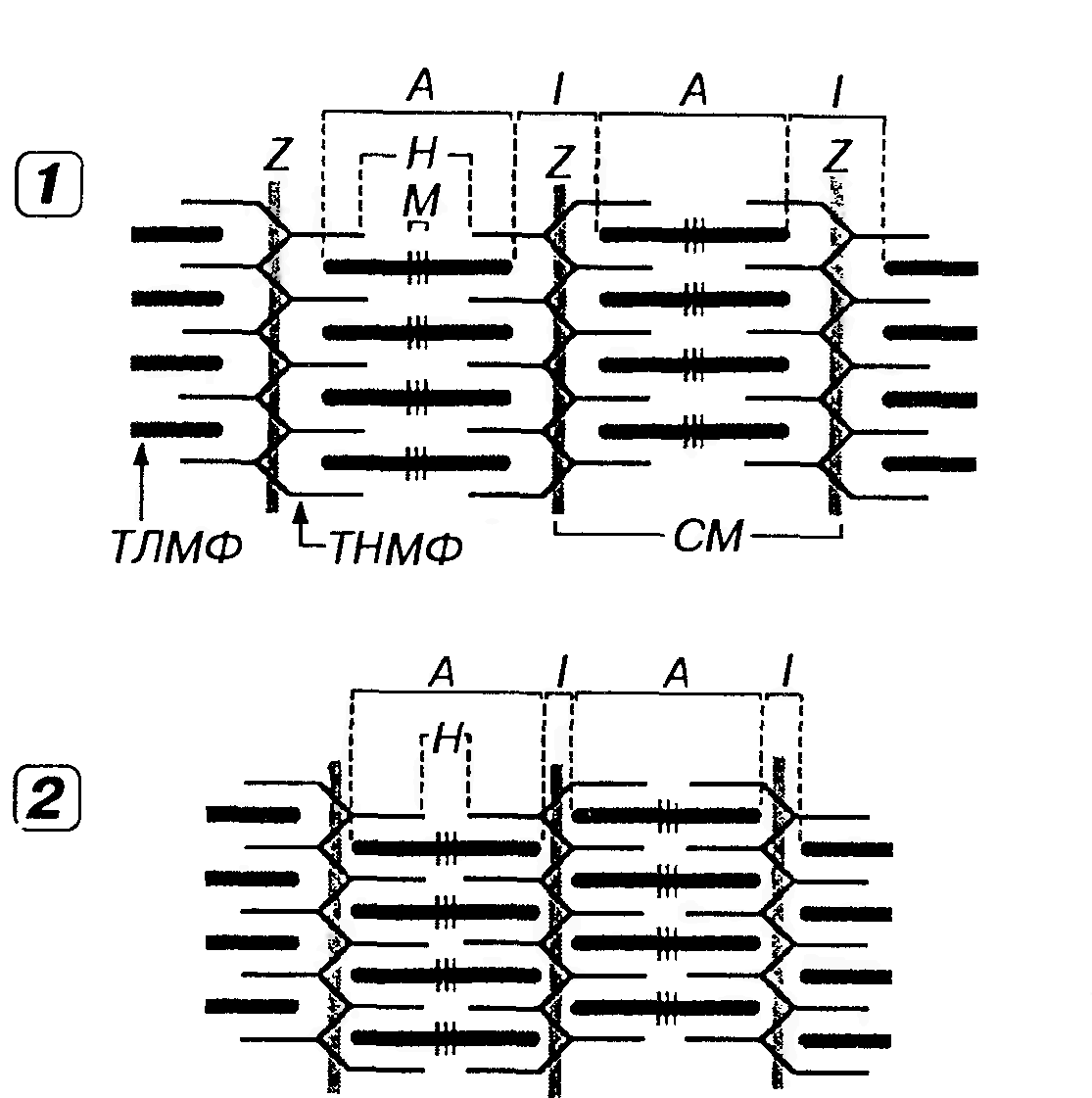
Сократительный аппарат мышечного волокна представлен миофибриллами - специальными органеллами, которые располагаются продольно в центральной части саркоплазмы и отделяются друг от друга рядами вытянутых митохондрий и цистерн саркоплазматической сети. На поперечном разрезе волокна видно, что миофибриллы симпласта образуют особые группы - поля Конгейма (рис. 40), которые, по мнению ряда авторов, являются артефактом. Миофибриллы имеют вид нитей диаметром 1-2 мкм и длиной, сопоставимой с протяженностью волокна. Их количестве в отдельном волокне варьирует в широких пределах (от нескольких десятков до 2000 и более). Они обладают собственной поперечной исчерченностью, причем в мышечном волокне они располагаются столь упорядочение, что А- и I-диски одних миофибрилл точно совпадают с аналогичными дисками других, обусловливая поперечную исчерченность всего волокна. Структурно-функциональной единицей миофибриллы является саркомер (миомер). Саркомер (миомер) представляет собой участок миофибриллы, расположенный между двумя телофрагмами (Z -линиями) и включающий А-диск и две половины I-дисков - по одной половине с каждой стороны (рис. 41). В расслабленной мышце длина саркомера составляет около 2-3 мкм, а ширина его участков выражается соотношением Н: А: I = 1: 3: 2; при сокращении мышцы саркомер укорачивается до 1,5 мкм. Миофибрилла типичного мышечного волокна человека длиной около 5 см насчитывает порядка 20 тыс. последовательно расположенных саркомеров.
Структура саркомера представлена упорядоченной системой толстых и тонких белковых нитей (миофиламентов). Толстые нити (диаметром около 10-12 нм и длиной 1,5-1,6 мкм) связаны с мезофрагмой и сосредоточены в А-диске, а тонкие (диаметром 7-8 нм и длиной 1 мкм) прикреплены к телофрагмам, образуют I -диски и частично проникают в А-диски между толстыми нитями (более светлый участок А-диска, свободный от тонких волокон, называется полоской H). В саркомере насчитывается несколько сотен толстых нитей. По сечению саркомера толстые и тонкие нити располагаются высокоорганизованно в узлах гексагональной решетки. Каждая толстая нить окружена шестью тонкими, каждая из тонких нитей частично входит в окружение трех соседних толстых (рис. 41). Толстые нити (миофиламенты) образованы упорядоченно упакованными молекулами фибриллярного белка миозина, на который приходится около 54% всех белков миофибриллы. Молекула миозина имеет вид нити длиной 150 нм и толщиной 2 нм. На одном из концов эта молекула содержит две округлые головки длиной около 20 нм и шириной около 4 нм (рис. 42). Протеолитическими ферментами миозин расщепляется на две фракции - легкий меромиозин ("стержень" молекулы миозина) и тяжелый меромиозин (участки головок и шейки, связывающие их со стержневой частью). Молекула миозина может сгибаться, как на шарнирах, в месте соединения тяжелого меромиозина с легким и в области прикрепления головки. Стержневые части молекул миозина собраны в пучки (численностью до 200 и более). Такие пучки, соединенные зеркально концами друг с другом в области М-линии, формируют толстые нити с центральной гладкой частью длиной около 0,2 мкм и двумя периферическими участками, в которых от центрального стержня отходят миозиновые головки (около 500). Миозин головок обладает АТФазной активностью (способностью осуществлять гидролиз АТФ), однако в отсутствие его взаимодействия с актином скорость гидролиза АТФ ничтожно мала.
Рис. 42. Строение толстых миофиламентов (по К.Де Дюву, 1987, с изменениями). 1 - молекула миозина имеет вид нити с двумя головками (Г) на одном конце. Миозин включает легкий меромиозин (ЛММ), образующий стержневую часть молекулы, и тяжелый меромиозин (ТММ), соответствующий участкам Г и связующей шейки. Участки сгибания молекулы миозина показаны стрелками. 2 - стержневые части молекул миозина собраны в пучки (П), снаружи которых располагаются миозиновые Г. 3 - толстые миофиламенты (ТЛМФ) образованы П молекул миозина, соединенными зеркально концами друг с другом. Центральная часть ТЛМФ - гладкая, периферические содержат многочисленные миозиновые Г.
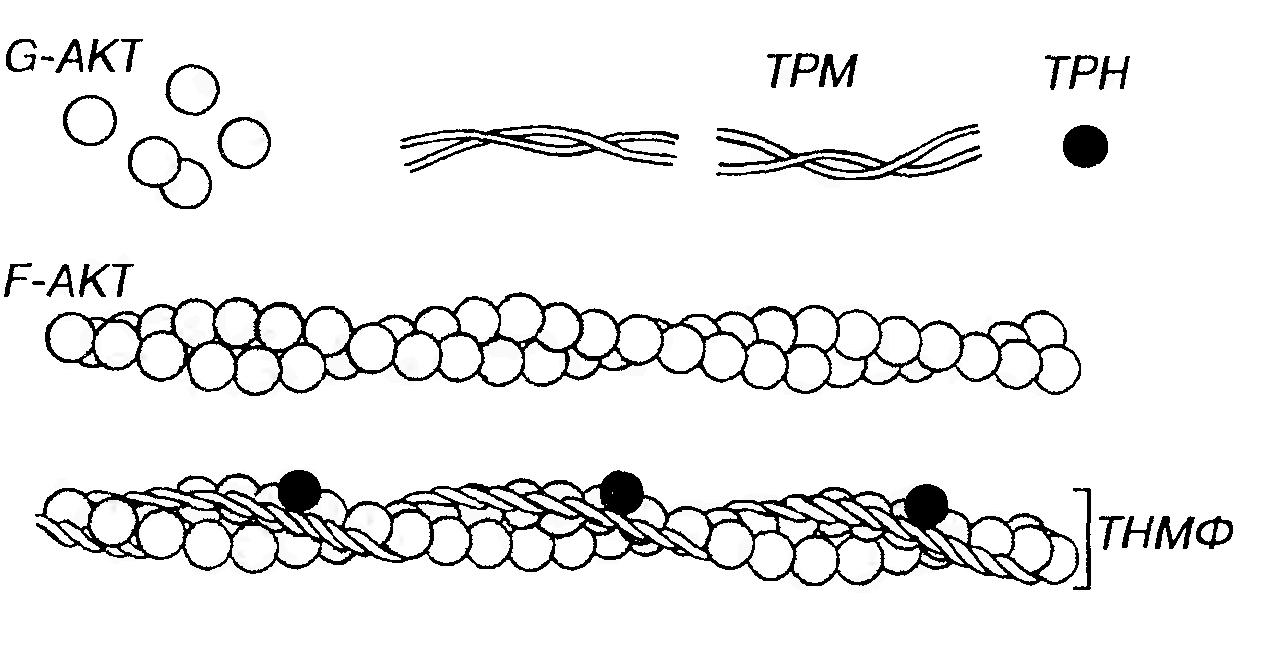
Тонкие нити (миофиламенты) содержат сократимый белок актин (на него приходится 20% белков миофибриллы) и два регуляторных белка - тропонин (около 2%) и тропомиозин (около 7%). Последние формируют функционально единый тропонин-тропомиозиновый комплекс. Актин в мономерной форме представлен полярными глобулярными субъединицами диаметром 4-5 нм (G -актин), которые имеют активные центры, способные связываться с молекулами миозина. G-актин агрегирует с образованием полимерного фибриллярного актина (F -актина), молекула которого имеет вид двух скрученных нитей толщиной 7 нм и вариабельной длины (рис. 43).
Рис. 43. Строение тонких миофиламентов. Тонкие миофиламенты (ТНМФ) содержат сократимый белок актин (АКТ) и два регуляторных белка - тропонин (TPH) и тропомиозин (ТРМ). Глобулярные субъединицы АКТ (G - AKT), агрегируют с образованием фибриллярного АКТ (F - AKT), молекула которого имеет вид двух скрученных нитей. ТРМ образован нитевидными молекулами, соединяющимися своими концами и образующими тяж, лежащий в борозде молекулы F - AKT. TPH - глобулярный белок, связанный с молекулой ТРМ и формирующий с ней функционально единый комплекс ТРН-ТМ.
Тропомиозин представлен нитевидными молекулами, которые соединяются своими концами, образуя длинный тонкий тяж, лежащий в борозде, образуемой перевитыми нитями F-актина. Так как таких борозд на молекуле актина две, то и тропомиозиновых нити тоже две. Всего в состав тонкой нити входит примерно 50 молекул тропомиозина.
Тропонин представляет собой глобулярный белок, каждая его молекула располагается на тропомиозиновой молекуле вблизи ее конца. Тропонин состоит из трех субъединиц: ТnС - связывающей кальций, ТnТ - прикрепляющейся к тропомиозину, и TnI - ингибирующей связывание миозина с актином. Механизм мышечного сокращения описывается теорией скользящих нитей, согласно которой укорочение каждого саркомера (а, следовательно, миофибрилл и всего мышечного волокна) при сокращении происходит благодаря тому, что тонкие нити вдвигаются в промежутки между толстыми без изменения их длины (рис. 44). Скольжение нитей в саркомере и усилие, развиваемое мышцей, обеспечиваются благодаря циклической активности миозиновых мостиков, которые при сокращении повторно прикрепляются к актину, обеспечивают усилие тяги, а затем открепляются от него (рис. 45). В этом механизме АТФ играет двойную роль, обеспечивая энергию, необходимую как для осуществления сокращения, так и для открепления мостков.
Рис. 44. Механизм мышечного сокращения в соответствии с теорией скользящих нитей. Укорочение саркомеров (СМ) при сокращении (2) по сравнению с их состоянием в покое (1) происходит благодаря тому, что тонкие миофиламенты (ТНМФ) вдвигаются в промежутки между толстыми (ТЛМФ) без изменения их длины. Остальные обозначения - как на рис. 41.
Строгая пространственная упорядоченность взаимодействия множества толстых и тонких нитей в саркомере определяется наличием сложно организованного поддерживающего аппарата. Его элементы на всех этапах мышечного сокращения и расслабления, динамично перестраиваясь, фиксируют и удерживают миофиламенты в правильном положении, которое оптимальным образом обеспечивает их взаимный контакт, взаимодействие и взаимное скольжение.
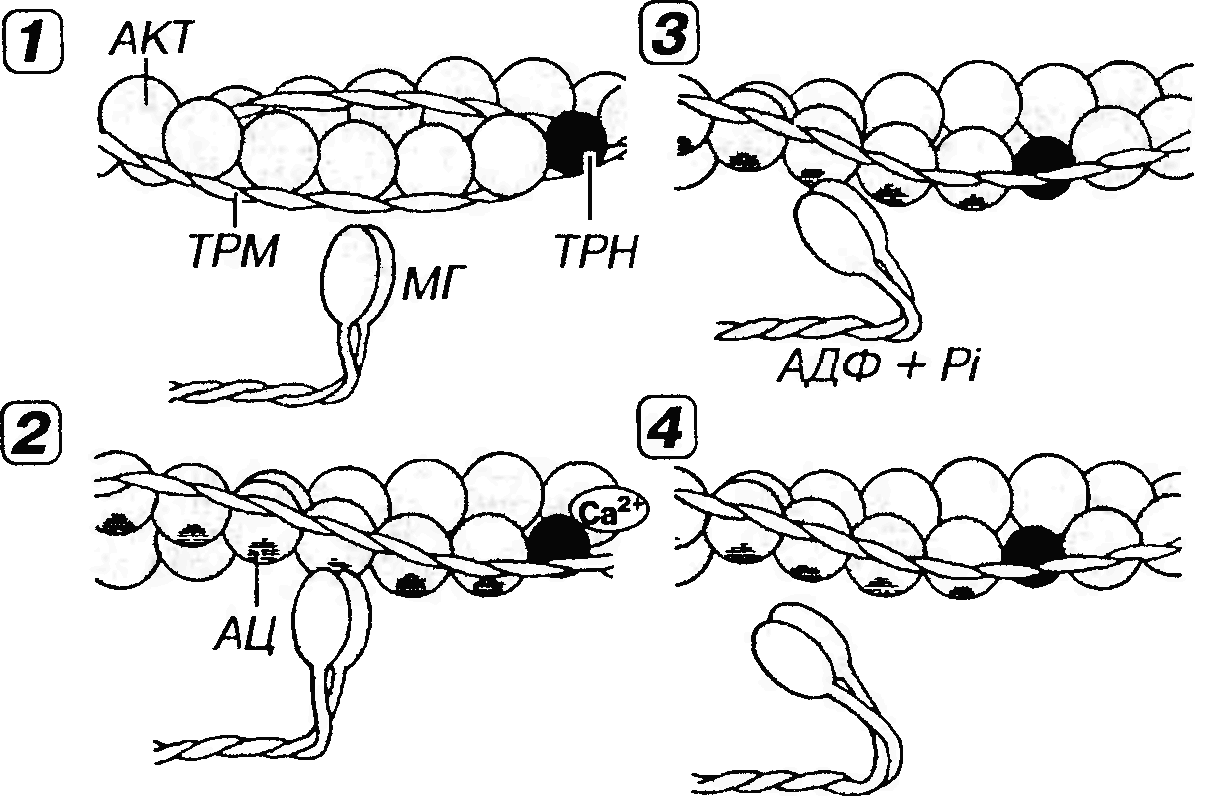
Рис. 45. Молекулярные механизмы мышечного сокращения. 1 - в покое миозиновые головки (МГ), с которыми связаны молекулы АТФ, неспособны взаимодействовать с активными центрами (АЦ) на молекуле актина (АКТ), потому что последние прикрыты комплексом тропонин-тропомиозин (ТРН-ТРМ). 2 - мышечное сокращение начинается вследствие повышения концентрации Са2+, который воздействует на ТРН. Возникающее изменение конформации ТРН и смещение молекулы связанного с ним ТРМ демаскирует АЦ на молекуле АКТ, с которыми связываются МГ, образуя поперечные мостики. 3 - за счет сгибания МГ в области их прикрепления к молекуле АКТ развивается усилие, смещающее тонкие миофиламенты (ТНМФ) вдоль толстых (ТЛМФ) к центру саркомера (см. рис. 44). АТФ при этом гидролизуется до АДФ и фосфата (Pi). 4 - размыкание мостика и его отделение от ТНМФ наступают вследствие связывания с ним новой молекулы АТФ. Далее мостик принимает исходное положение (перпендикулярное ТНМФ) и начинается новый цикл сокращения. Циклическое взаимодействие МГ и ТНМФ будет продолжаться при сохранении высокой концентрации ионов Са2+ и наличии АТФ.
В покое (при очень низкой концентрации ионов Са2+) в миофибрилле расслабленного мышечного волокна толстые и тонкие нити не соприкасаются. Миозиновые головки (с которыми связаны молекулы АТФ) не могут взаимодействовать с активными центрами (участками связывания миозина) на молекуле актина, потому что последние прикрыты тропонин-тропомиозиновым комплексом. Толстые и тонкие филаменты беспрепятственно скользят друг относительно друга. При этом мышечные волокна почти не сопротивляются пассивному растяжению. Такое состояние свойственно разгибательной мышце при сокращении соответствующей сгибательной. В отсутствие тропомиозина и тропонина (в условиях in vitro) миозин непрерывно взаимодействует с актином (пока имеется АТФ).
Мышечное сокращение вызывается резким повышением концентрации ионов Са2+ в области миофиламентов и включает несколько этапов (см. рис. 45 [2-4]). A. Связывание ионов Са2+ с тропонином и освобождение активных центров на молекуле актина. Ионы Са2+ связываются с ТnС-субъединицами тропонина на тонких филаментах. При этом тропонин изменяет свою конформацию, смещает молекулы тропомиозина и открывает активные центры (участки связывания миозина) на молекуле актина. Б. Связывание миозина и актина (формирование поперечных мостиков). Миозиновые головки связываются с активными центрами на молекуле актина, формируя мостики, расположенные перпендикулярно продольной оси нити. Менее чем через 1 мс после этого под влиянием актомиозинового комплекса происходит гидролиз АТФ и отщепление его продуктов (АДФ и неорганического фосфата). При этом угол наклона мостика относительно продольной оси нити изменяется до 40°. Такой конформационный переход, происходящий в области прикрепления головки миозиновой молекулы, обусловливает развитие усилия и смещение тонких филаментов к центру саркомера. Предполагается, что "рабочий ход" миозинового мостика составляет около 10 нм; таким образом, за один цикл мостик вызывает относительное перемещение тонких нитей на расстояние, равное примерно 1/200 длины саркомера. B. Размыкание мостика. Связывание новой молекулы АТФ с мостиком вызывает его отделение от тонкого филамента. Мостик размыкается, возвращаясь в прежнее положение относительно миозиновой нити и может прийти в замыкание со следующим активным центром на тонкой. Каждый цикл замыкания-размыкания сопровождается расщеплением молекулы АТФ. В живой мышце это осуществляется с интервалом в несколько десятков миллисекунд после присоединения новой молекулы АТФ. В трупной мышце, где АТФ отсутствует, мостик не может разомкнуться, и мышца переходит в состояние трупного окоченения (rigor mortis). При сокращении мышцы не происходит одновременного замыкания всех мостиков - их число нарастает по ходу его развития. При последующем расслаблении мышцы число мостиков снижается. Изменение длины саркомера при сокращении является результатом относительного продольного смещения толстых и тонких нитей. При этом ширина А-диска не меняется; по мере проникновения в него тонких нитей происходит укорочение I-диска; соответственно значительно сужается Н-полоска (рис. 44). Расслабление после мышечного сокращения происходит в результате снижения концентрации Са2+ в области саркомера, которое вызывает отщепление Са2+ от TnC-субъединицы тропонина и возвращение тропонина в первоначальное конформационное состояние. Нити тропомиозина при этом вновь закрывают активные центры на молекулах актина, что обусловливает прекращение циклического образования мостиков. Аппарат передачи возбуждения (саркотубулярная система) необходим для того, чтобы распространяющаяся по сарколемме волна деполяризации могла вызвать срабатывание сократительного аппарата миофибрилл. В мышечном волокне связь между возбуждением и сокращением выполняют две специализированные мембранные системы - саркоплазматическоя сеть и поперечные (Т-) трубочки (от англ. transverse - поперечный), образующие функционально единую саркотубулярную систему (рис. 46).
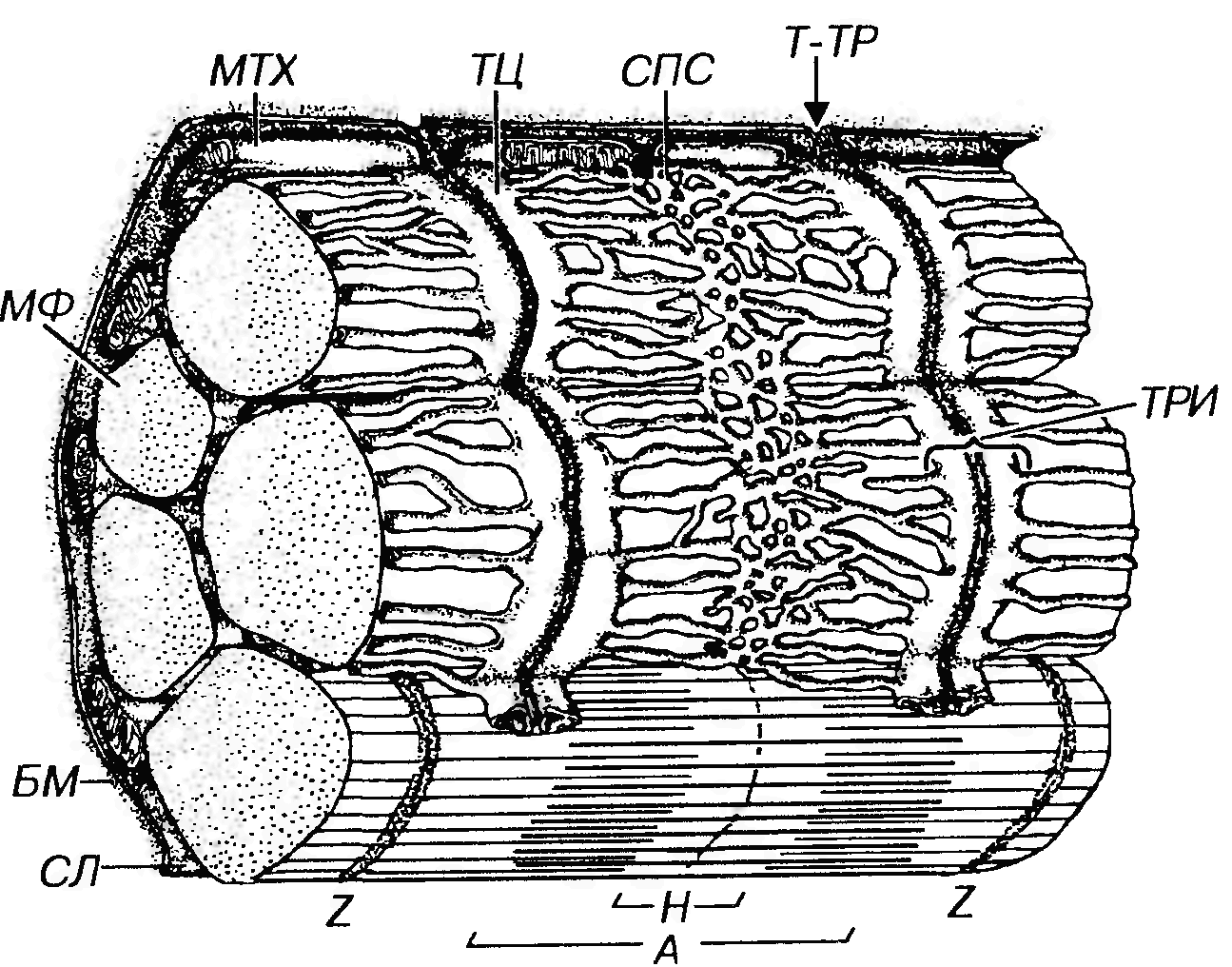
Рис. 46. Саркотубулярная система волокна скелетной (соматической) мышечной ткани. Саркотубулярная система включает саркоплазматическую сеть (СПС) и поперечные, или Т-трубочки (Т-ТР). СПС окружает каждый саркомер миофибриллы; ее трубочки сливаются, образуя пары плоских терминальных цистерн (ТЦ). T - TP представляют собой впячивания сарколеммы (СЛ), отходящие от нее под прямым углом и проникающие в промежуток между двумя ТЦ, в совокупности с которыми они формируют триады (ТРИ). МФ - миофибриллы. Обозначения компонентов саркомера - те же, что на рис. 41.
Саркоплазматическоя сеть - система уплощенных, вытянутых и анастомозирующих мембранных трубочек и мешочков, которая окружает каждый саркомер миофибриллы наподобие муфты. В области наружных отделов А- и I-дисков трубочки сливаются, образуя пары плоских терминальных цистерн (на каждый саркомер приходится по две такие пары). Саркоплазматическая сеть обладает выраженной способностью депонировать и выделять ионы кальция. Ее мембрана содержит высокие концентрации интегральных белков, являющихся кальциевыми насосами, а на внутренней поверхности находится белок кальсеквестрин, связывающий ионы Са2+. Поперечные (Т-) трубочки представляют собой впячивания сарколеммы, отходящие от нее под прямым углом к оси волокна и расположенные у млекопитающих вблизи границы I- и А- дисков. Ветви соседних Т-трубочек опоясывают каждый саркомер и анастомозируют друг с другом. Конечные участки Т-трубочек проникают в промежуток между двумя терминальными цистернами саркогшазматической сети (рис. 46), формируя вместе с ними особые структуры - триады. В области триады между параллельно лежащими мембранами Т-трубочки и терминальных цистерн, разделенными узкой щелью, имеются специализированные контакты, которые образованы рядами плотных частиц (ножек), предположительно служащие каналами выделения кальция. Выделение кальция происходит после того, как волна деполяризации с поверхности сарколеммы по Т-трубочкам распространяется вглубь волокна. В области триад возбуждение передается на мембрану саркоплазматической сети и вызывает повышение ее проницаемости. Это приводит к быстрому выделению из ее элементов ионов кальция (главным образом, в области терминальных цистерн). Выделившийся Са2+ диффундирует в миофибриллы, где он, присоединяясь к тропони-ну, запускает механизм взаимодействия актина и миозина (см. выше). Концентрация Са2+ вокруг миофиламентов при этом резко повышается с 10-7М до 10-5М. Активный обратный транспорт кальция в саркоплазматическую сеть (секвестрация кальция) происходит наряду с его выбросом, который представляет собой кратковременный процесс. Обратный транспорт Са2+ осуществляется благодаря деятельности кальциевых насосов (Са-зависимой АТФазы) в мембране саркоплазматической сети. Падение концентрации Са2+ вследствие его секвестрации приводит к возвращению тропонина в первоначальное конформационное состояние, прекращению взаимодействия миозиновых мостиков с актином и расслаблению мышечного волокна. Опорный аппарат мышечного волокна включает особые элементы цитоскелета, обеспечивающие высокоупорядоченное расположение миофиламентов и миофибрилл внутри волокна, а также связанную с ними сарколемму и базальную мембрану (см. рис. 41, 45 и 46), соединяющие мышечное волокно с сухожилием, на которое передается усилие, развиваемое волокном при сокращении. Телофрагма (Z -линия) - область прикрепления тонких миофиламентов двух соседних саркомеров; она имеет вид плотной полоски шириной 30-100 нм без резких границ. Представляет собой сложную трехмерную решетку из особых тонких нитей (Z -филаментов), идущих зигзагообразно под углом 45° к оси саркомера и образующих тетрагональную (четырехугольную) структуру, связывающую тонкие нити двух соседних саркомеров. В ячейках решетки этих филаментов имеется плотный материал. В состав Z-линий входит ряд белков: α-актинин, филамин, Z-белок. Мезофрагма (М-линия) - плотная линия шириной 75-85 нм, расположенная в центре А-диска и являющаяся областью закрепления толстых (миозиновых) филаментов в саркомере. Она образована центральными участками миозиновых филаментов, которые располагаются в виде гексагональных фигур и связаны друг с другом системой мостиков, состоящих из тонких нитей белков миомезина, креатинкиназы и М-белка. Титин (коннектин) представляет собой белок с эластическими свойствами, нити которого присоединены к толстым филаментам по всей их длине и, продолжаясь в I-диски, прикрепляют концы толстых филаментов к Z-линиям. Таким образом, нити титана связывают М- и Z-линии, и, благодаря своей эластичности, препятствуют перерастяжению мышцы. Они образуют внутри саркомера решетчатую структуру и поддерживают упорядоченное взаимное расположение системы толстых и тонких миофиламентов. Небулин - белок, имеющий вид удлиненных нитей, расположенных по всей ширине I-диска параллельно тонким филаментам, с которыми он связан. Предполагается, что небулин отвечает за поддержание длины тонких филаментов и (или) обеспечивает их механическую стабилизацию. Промежуточные филаменты (диаметром около 10 нм), состоящие из белка десмина, являются важным элементом цитоскелета и образуют в пределах мышечного волокна две пространственные системы. Первая состоит из филаментов, которые располагаются в саркомерах продольно и связывают соседние телофрагмы одной миофибриллы. Вторая представлена поперечно ориентированными филаментами, которые связывают мезофрагмы, а также телофрагмы соседних миофибрилл друг с другом. Такие же филаменты прикрепляют телофрагмы к сарколемме и элементам системы Т-трубочек и саркоплазматической сети. Благодаря описанной организации системы промежуточных филаментов поддерживается упорядоченное взаимное расположение саркомеров соседних миофибрилл и других компонентов мышечного волокна. Дистрофин - белок, одними участками прикрепляющийся к актиновым филаментам, а другими - к комплексу гликопротеинов, которые пронизывают сарколемму и связываются на ее поверхности с компонентами базальной мембраны. Таким путем усилия, создаваемые внутри мышечного волокна, посредством ряда белков передаются на элементы межклеточного вещества. Генетический дефект, связанный с нарушением выработки дистрофина, обусловливает развитие мышечного заболевания - дистрофии Дюшенна. Костамеры - кольца из белка винкулина, охватывающие изнутри мышечное волокно и расположенные перпендикулярно к его длинной оси. Они представляют собой участки непосредственного соединения между сарколеммой и подлежащими 1-дисками миофибрилл. Благодаря наличию в костамерах интегринов они, также, возможно, являются структурами, которые через адгезивный гликопротеин фибронектин связывают элементы межклеточного вещества (например, коллагеновые волокна) с миофибриллами. Помимо винкулина, в костамерах имеются другие белки, связанные с цитоскелетом: талин, спектрин, α-актинин. Структура краевых участков мышечных волокон. На концах мышечных волокон сарколемма, покрытая базальной мембраной, образует многочисленные глубокие впячивания, в которые вдаются коллагеновые волокна сухожилия, вплетающиеся в базальную мембрану и прочно связывающие сухожилие с мышечными волокнами. Энергетический аппарат мышечных волокон представлен митохондриями, вырабатывающими энергию, необходимую для осуществления мышечной работы, синтетических, транспортных и других процессов жизнеобеспечения, а также трофическими включениями, содержащими вещества, расщепление которых служит источником энергии. Митохондрии в миосимпласте располагаются в виде цепочек под сарколеммой и между миофибриллами (рис. 46). Они имеют вытянутую форму, содержат большое количество поперечно расположенных ламеллярных крист, характеризуются высокой активностью окислительно-восстановительных ферментов. Их содержание и размеры больше в красных волокнах, чем в белых и увеличиваются при тренировке мышц. Энергия, необходимая для осуществления мышечной работы, запасается в мышечных волокнах в виде АТФ и фосфокреатина - энергоемких фосфатных соединений. Источником энергии служит расщепление гликогена и липидов. При кратковременных резких нагрузках на скелетные мышцы источником энергии служит глюкоза, получаемая преимущественно в результате расщепления гликогена. Главным источником энергии при выполнении работы, требующей выносливости, служат жирные кислоты. Гликоген находится в саркоплазме (преимущественно белых волокон) в виде β-частиц диаметром 20-30 нм. Последние образуют скопления между миофибриллами, большей частью на уровне I-дисков. Запасы гликогена, составляющие 0,5-1% массы волокна, опустошаются при длительной интенсивной нагрузке. Липидные капли располагаются между миофибриллами по всей толщине миосимпласта, образуя скопления преимущественно на уровне I-дисков. Их содержание варьирует в широких пределах, но в среднем выше в красных волокнах (0,5% объема саркоплазмы), чем в белых (0,2%). Миоглобин - железосодержащий кислород-связывающий пигмент мышечных волокон, придающий им красный цвет и сходный по строению и функции с гемоглобином эритроцитов - типичное включение мышечного волокна, которое можно условно отнести к энергетическому аппарату. Миоглобин находится в более высоких концентрациях в красных волокнах (что и определяет их цвет); его способность к связыванию кислорода способствует повышению активности процессов окислительного фосфорилирования). Синтетический аппарат мышечного волокна представлен свободными рибосомами и полирибосомами (особенно многочисленными под сарколеммой в области I-диска и вблизи ядер), цистернами грЭПС и комплексом Гольджи, элементы которого в виде сотен или тысяч стопок мешочков рассеяны по саркоплазме миосимпласта. Лизосомальный аппарат (аппарат внутриклеточного переваривания) в мышечных волокнах необходим для обеспечения постоянно протекающего процесса обновления его структурных компонентов. Содержание лизосом связано с функциональной активностью мышцы и возрастом человека. Остаточные тельца лизосомального генеза, содержащие липофусцин, становятся многочисленными при старении и, в особенности, при резком снижении функциональной активности мышцы. Миосателлитоциты - мелкие уплощенные клетки, располагающиеся в неглубоких вдавлениях сарколеммы миосимпластической части мышечного волокна и покрытые вместе с ней общей базальной мембраной (рис. 41). Ядро миосателлитоцита - плотное, относительно крупное (занимает почти всю клетку), с более высоким содержанием гетерохроматина, чем в ядрах миосимпласта, органеллы мелкие и немногочисленные. Эти клетки представляют собой камбиальные элементы скелетной мышечной ткани. Они активируются при повреждении мышечных волокон и обеспечивают их репаративную регенерацию. Сливаясь с симпластической частью волокна при усиленной нагрузке, миосателлитоциты участвуют в его гипертрофии. В мышечных волокнах у плода и новорожденного доля ядер миосателлитоцитов достигает 30-35% от общего содержания ядер; после рождения она быстро снижается, составляя в детстве 7-10%, а у взрослого - около 5%. Содержание этих клеток выше красных волокнах, чем в белых. Типы мышечных волокон Мышечные волокна в скелетных мышцах позвоночных животных и человека обладают, несмотря на общий план строения, определенными структурными, биохимическими и функциональными различиями. Используемые классификации мышечных волокон основаны на учете их различных признаков и совпадают неполностью. В обобщенном виде можно условно выделить три основных типа мышечных волокон, между которыми существуют переходные варианты: тип 1 (красные), тип IIB(белые) и тип IIA (промежуточные). Тип I - красные, медленные, тонические, устойчивые к утомлению, с небольшой силой сокращения, окислительные. Характеризуются малым диаметром, относительно тонкими миофибриллами, высокой активностью окислительных ферментов (например, сукцинатдегидрогеназы - СДГ), низкой активностью гликолитических ферментов и миозиновой АТФазы, преобладанием аэробных процессов, высоким содержанием миоглобина (определяющим их красный цвет), крупных митохондрий (занимают около 15% объема саркоплазмы) с многочисленными кристами и липидных включений, широкой (50-100 нм) Z-линией, высоким содержанием миосателлитоцитов, богатым кровоснабжением. Численно преобладают в мышцах, выполняющих длительные тонические нагрузки. Тип II В - белые, быстрые, тетанические, легко утомляющиеся, с большой силой сокращения, гликолитические. Характеризуются большим диаметром, крупными и сильными миофибриллами, высокой активностью гликолитических ферментов (например, лактатдегидрогеназы - ЛДГ) и АТФазы, низкой активностью окислительных ферментов, преобладанием анаэробных процессов, относительно низким содержанием митохондрий (более мелких и с менее развитыми кристами, чем в волокнах I типа и занимающих около 7% объема саркоплазмы), липидов и миоглобина (определяющим их светлый цвет), значительным количеством гликогена, узкой (30-40 нм) Z-линией, относительно небольшим числом миосателлитоцитов, сравнительно слабым кровоснабжением. Преобладают в мышцах, выполняющих быстрые движения, например, мышцах конечностей.
Тип II А - промежуточные, быстрые, устойчивые к утомлению, с большой силой, оксилителъно-гликолитические. На препаратах напоминают волокна типа I. В равной степени способны использовать энергию, получаемую путем окислительных и гликолитических реакций. По своим морфологическим и функциональным характеристикам занимают положение, промежуточное между волокнами типа I и IIВ. Красные и белые волокна различаются также содержанием различных изоформ миозина и субъединиц тропонина. В частности, изоформы миозина, характерные для белых волокон, отличаются более быстрой циклической активностью миозиновых мостиков, а, следовательно, большей скоростью сокращения. Соотношение числа волокон различных типов в мышце. Скелетные мышцы человека являются смешанными, т.е. содержат волокна различных типов, которые распределены в них мозаично. Соотношение красных и белых волокон в мышцах каждого человека индивидуально, предопределено генетически и почти не меняется с возрастом. В мышцах большинства людей белые и красные волокна содержатся примерно в равных количествах. Вместе с тем, у отдельных людей могут преобладать волокна того или иного типа, что позволяет им более успешно справляться с длительной физической нагрузкой небольшой мощности или с кратковременной тяжелой нагрузкой. Изменения в волокнах различных типов при тренировке мышц неодинаковы и зависят от характера нагрузок. Нарастание массы мышц при этом связано с увеличением диаметра (гипертрофией) мышечных волокон (главным образом, белых); в последние годы вновь высказываются взгляды о возможности некоторого увеличения и числа волокон при очень высоких нагрузках.
Физиологическая регенерация волокон скелетной мышечной ткани непрерывно осуществляется в нормальных условиях на ультраструктурном уровне и состоит в самообновлении их органелл и других структурных компонентов, обеспечивающем поддержание баланса между анаболическими и катаболическими процессами.
Гипертрофия мышечных волокон развивается в ответ на повышенные нагрузки в результате преобладания анаболических процессов над катаболическими. Она проявляется увеличением содержания компонентов их саркоплазмы; при этом нагрузки, требующие выносливости, вызывают увеличение всего объема саркоплазмы и, особенно, митохондрий, а скоростно-силовые нагрузки - преимущественное нарастание массы миофибрилл (вследствие увеличения их числа и диаметра). Атрофия мышечных волокон возникает вследствие бездействия (при денервации или гипокинезии), а также при голодании. Денервация вызывает снижение массы мышцы на 50% и более, уменьшение диаметра волокон, дезорганизацию сократительного аппарата и элементов цитоскелета, сглаживание различий их типов. Наиболее быстро атрофируются белые волокна; красные изменяются в меньшей степени. Гипокинезия обусловливает более выраженные изменения в красных волокнах, которые более чувствительны к снижению нагрузки, чем белые, которые вовлекаются в процесс атрофии позднее. Выраженные явления мышечной атрофии развивается у космонавтов; наибольшие изменения при этом отмечены в красных мышечных волокнах. Голодание сопровождается распадом белков миофибриллярного аппарата и поражает в первую очередь белые волокна.
Репаративная регенерация мышечных волокон направлена на восстановление их целостности после повреждения и частично напоминает эмбриональный миогенез. При любых видах травмы процесс регенерации включает закономерную последовательность явлений: (1) инфильтрацию области повреждения фагоцитами, (2) восстановление целостности сосудов (реваскуляризацию), (3) фагоцитоз некротизированных мышечных волокон, (4) пролиферацию миогенных клеток-предшественников, (5) их последующее слияние с образованием мышечных трубочек, (6) дифференцировку трубочек с образованием зрелых мышечных волокон, (7) восстановление иннервации.
Миграция фагоцитов (нейтрофильных гранулоцитов и макрофагов) в область повреждения происходит под хемотаксическим действием продуктов, выделяемых травмированными волокнами. Устремляясь к поврежденным волокнам, фагоциты активно поглощают тканевой детрит, часто сохраняя базальную мембрану разрушенны
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-02-07; просмотров: 437; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.84.32 (0.091 с.) |