Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Рыхлая волокнистая соединительная тканьСодержание книги
Похожие статьи вашей тематики
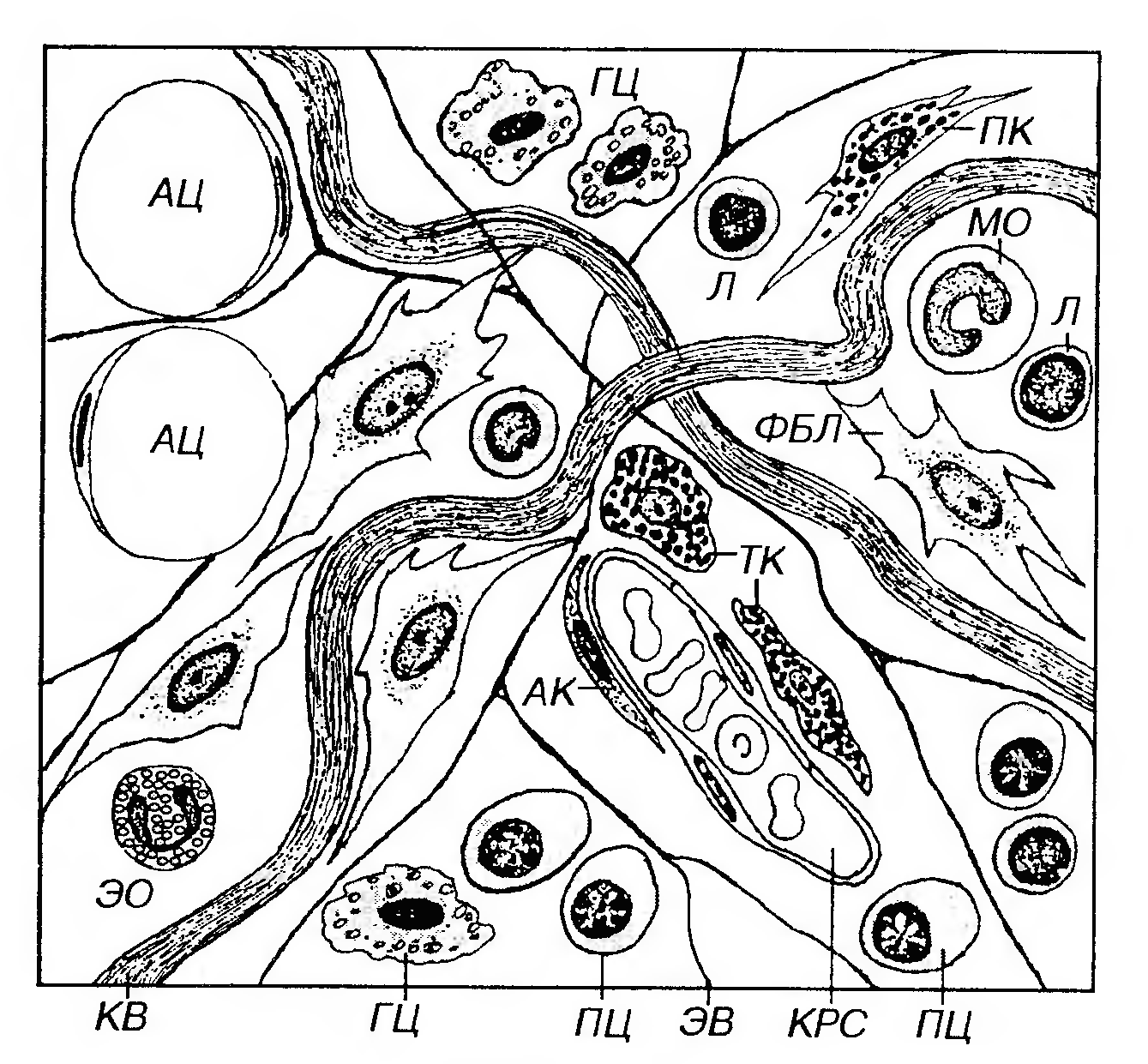
Поиск на нашем сайте Рыхлая волокнистая соединительная ткань является самым распространенным видом соединительных тканей и имеет наиболее типичное для этих тканей строение, так как содержит разнообразные клетки и все компоненты межклеточного вещества (рис. 35). Она выполняет все функции, свойственные соединительным тканям, взаимодействуя с другими тканями, связывая их между собой (что оправдывает общее название этой группы тканей) и способствуя поддержанию гомеостаза в организме. Эта ткань обнаруживается повсеместно, во всех органах - она образует их строму (основу), в частности, междольковые прослойки и прослойки между слоями и оболочками, заполняет пространства между функциональными элементами других тканей, сопровождает нервы и сосуды, входит всостав кожи и слизистых оболочек. Клетки рыхлой волокнистой соединительной ткани представляют собой сложную гетерогенную популяцию функционально разнообразных и взаимодействующих между собою и с компонентами межклеточного вещества элементов (рис. 35), которые условно объединяют в несколько групп.
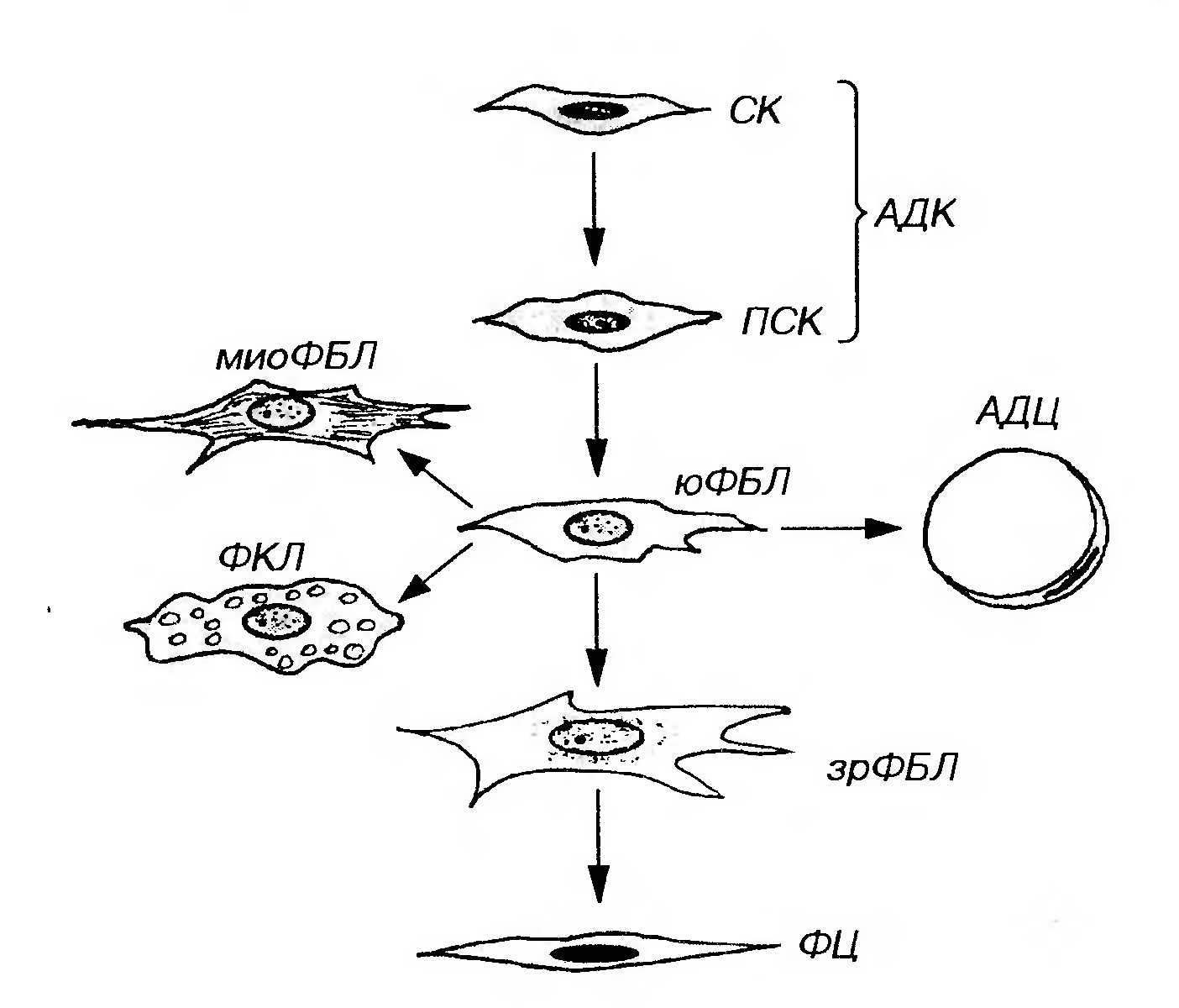
Рис. 35. Рыхлая волокнистая соединительная ткань. АК - адвентициальная клетка, КРС - кровеносный сосуд, ФБЛ - фибробласт, АЦ - адипоцит, ГЦ - гистиоцит, ПЦ - плазмоцит, ТК - тучная клетка, Л - лимфоцит, МО - моноцит, ЭО - эозинофил, ПК - пигментная клетка, KB - коллагеновые волокна, ЭВ - эластические волокна. По признаку постоянства присутствия в составе рыхлой волокнистой соединительной ткани ее клетки подразделяют на: (1) оседлые (фиксированные) клетки, т.е. образующиеся и постоянно пребывающие в этой ткани. К этой группе относят адвентициальные клетки, фибробласты, фиброциты и жировые клетки (адипопиты). В зрелой рыхлой волокнистой соединительной ткани содержание оседлых клеток относительно стабильно; (2) блуждающие клетки (иммигранты) - подвижные элементы, поступающие в соединительную ткань из крови. В эту группу включают все виды лейкоцитов (гранулоцитов и агранулоцитов). Содержание этих клеток в отдельных участках соединительной ткани может существенно изменяться при различных иммунных реакциях и воспалении. Макрофаги (гистиоциты), плазматические и тучные клетки одни авторы считают оседлыми элементами (поскольку они образуются в соединительной ткани и постоянно присутствуют в ней), другие причисляют к блуждающим клеткам (так как они дифференцируются из предшественников, циркулирующих в крови). Фибробласты (от лат. fibra - волокно и греч. blastos - росток) ̶ наиболее распространенные и функционально ведущие клетки рыхлой волокнистой соединительной ткани. Функции фибробластов: 1) продукция всех компонентовмежклеточного вещества (волокон и основного аморфного вещества); 2) поддержание структурной организации и химического гомеостаза межклеточного вещества (за счет сбалансированных процессов его выработки и разрушения);3) регуляция деятельности других клетоксоединительных тканей и влияние на другие ткани. Развитие фибробластов. Источником развития фибробластов в эмбриогенезе является мезенхима. После рождения фибробласты представляют собой сложную систему (дифферон) клеток, имеющих общего предшественника и различающихся по степени дифференцировки, морфологическим и функциональным характеристикам. Основная линия развития в этом диффероне (рис. 36) представлена последовательностью: стволовая клетка → полустволовая клетка- предшественник → малодифференцированный (юный) фибробласт → зрелый (дифференцированный) фибробласт → фиброцит.
Рис. 36. Дифферон фибробластов. СК - стволовая клетка, ПСК - полустволовая клетка-предшественник, АДК - адвентициальная клетка, юФБЛ - юный (малодифференцированный) фибробласт, зрФБЛ - зрелый (дифференцированный) фибробласт, ФЦ - фиброцит, АДЦ - адипоцит, ФКЛ - фиброкласт, миоФБЛ - миофибробласт. Стволовая клетка и полустволовые клетки-предшественники, образующиеся из нее в ходе дифференцировки, представляют собой наиболее ранние элементы дифферона фибробластов. Морфологически им, по всей видимости, соответствует адвентициальная клетка ̶ мелкая веретеновидная уплощенная малодифференцированная клетка, располагающаяся по ходу капилляров (см. рис. 35). Для нее характерно темное ядро и базофильная цитоплазма, содержащая слабо развитые органеллы. Стволовые клетки устойчивы к повреждающим воздействиям, редко делятся и образуют самоподдерживающуюся популяцию. Полустволовые клетки при стимуляции способны к высокой митотической активности, однако их синтетический аппарат не развит, и они не продуцируют компонентов межклеточного вещества соединительной ткани. Вопрос о природе и свойствах стволовой клетки окончательно не разработан. Малодифференцированный (юный) фибробласт - базофильная клетка более крупных размеров, чем адвентициальная, с небольшим числом отростков. Для нее характерно крупное круглое или овальное ядро с 1-2 ядрышками, умеренно развитый синтетический аппарат. Она сохраняет способность к пролиферации, но они уже начинает осуществлять синтез типичных компонентов межклеточного вещества соединительной ткани - коллагена и гликозаминогликанов. Способность юных фибробластов к направленной миграции определяет их важную роль в репаративных процессах, в частности, в заживлении ран. Миграция осуществляется благодаря наличию в их цитоплазме сократимых микрофиламентов, на которые опосредованно передаются сигналы с многочисленных рецепторов плазмолеммы, воспринимающих молекулы хемотаксических веществ. Факторами, привлекающими их в очаг повреждения, служат продукты, выделяемые макрофагами, Т-лимфоцитами, тромбоцитами,фибронектин, а также пептиды, образующиеся при расщеплении коллагена. Многие из этих факторов оказывают на юные фибробласты также митогенное действие, стимулируют их функциональную активность и дифференцировку, по завершении которой эти клетки превращаются в зрелые фибробласты. Зрелый (дифференцированный) фибробласт - крупная (на пленочных препаратах - более 40-50 мкм в поперечнике) отростчатая клетка с нерезкими границами и светлым ядром, содержащим мелкодисперсный хроматин и 1-2 ядрышка (см. рис. 35 и 37). Цитоплазма слабо базофильна и характеризуется диплазматшеской дифференцировкой - нерезким разделением на внутреннюю, более плотную часть, окружающую ядро, - эндоплазму и периферическую, сравнительно светлую и образующую отростки - эктоплазму. Эндоплазма содержит большую часть органелл мощно развитого синтетического аппарата, а эктоплазма заполнена преимущественно элементами цитоскелета. Цистерны грЭПС часто растянуты, содержат мелкозернистый материал низкой электронной плотности. В цитоплазме располагаются также лизосомы, митохондрии, липидные капли и многочисленные пузырьки. Все элементы цитоскелета хорошо выражены. Фибробласт обладает подвижностью, способностью изменять свою форму и обратимо прикрепляться к другим клеткам и компонентам межклеточного вещества (волокнам).
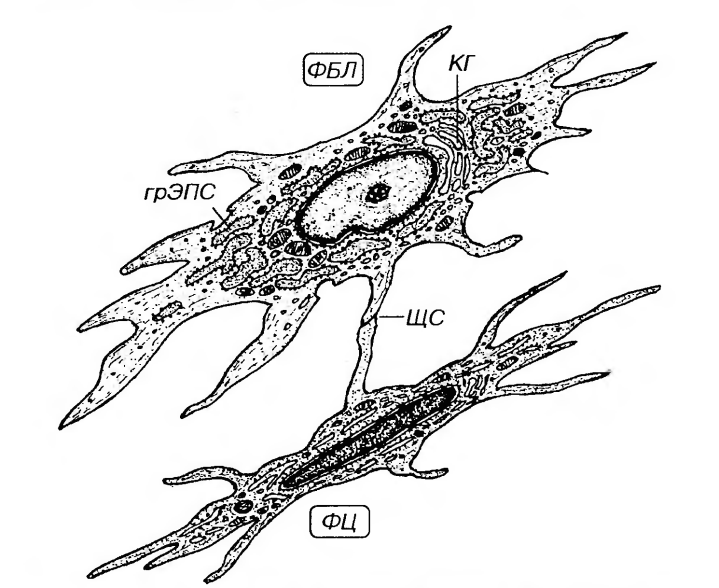
Рис. 37. Ультраструктурная организация фибробласта (ФБЛ) и фиброцита (ФЦ). ЩС - щелевое соединение (между отростками ФБЛ и ФЦ), КГ - комплекс Гольджи. Функции зрелого фибробласта заключаются в сбалансированных процессах продукции, перестройки и частичного разрушения межклеточного вещества, что обеспечивает возможность тонкой регуляции его архитектоники и состояния. Фибробласты оказывают также влияние на деятельность клеток других типов в соединительной и соседних с ней тканях. Регуляция деятельности фибробластов осуществляется факторами, вырабатываемыми макрофагами, Т-лимфоцитами, тромбоцитами и эпителиальными клетками (включая эндотелиоциты), а также различными гормонами. Регуляторное влияние фибробластов на другие клетки обеспечивается благодаря продукции ими гуморальных факторов, активно воздействующих на рост, дифференцировку и функциональную активность, как их собственной популяции, так и макрофагов, моноцитов, лимфоцитов, гладкомышечных и эпителиальных клеток. На указанные клетки в качестве локальных регуляторов воздействуют также вырабатываемые фибробластами компоненты межклеточного вещества (в особенности, фибронектин, гликозаминогликаны, коллагены различных типов). Большинство фибробластов разрушается в процессе жизнедеятельности, но часть их превращается в малоактивную долгоживущую форму - фиброциты. Фиброцит - конечная форма развития фибробласта - узкая веретенообразная, неспособная к пролиферации клетка с длинными тонкими отростками, которые часто имеют уплощенную крыловидную форму. Ядро - сравнительно плотное (с преобладанием гетерохроматина), занимает большую часть клетки. Цитоплазма содержит слабо развитый синтетический аппарат, значительное количество лизосом, липофусциновых гранул (рис. 37). Функция этих клеток состоит в регуляции метаболизма и поддержании стабильности межклеточного вещества; синтез его компонентов осуществляется ими очень слабо. Фиброциты располагаются между пучками коллагеновых волокон. Фиброкласты (от лат. fibra - волокно и греч. klasis - разрушение) - клетки дифферона фиброцитов, специализированные на функции разрушения межклеточного вещества соединительной ткани, которая резко преобладает над их синтетической и секреторной активностью. По-видимому, процессы деградации межклеточного вещества этими клетками осуществляются внутриклеточным и внеклеточным механизмами, аналогичными тем, что используются зрелыми фибробластами. В их цитоплазме выявляются многочисленные вакуоли, содержащие литические ферменты и коллагеновые фибриллы на различных стадиях лизиса. Эти клетки обеспечивают перестройку и инволюцию соединительной ткани; они особенно многочисленны в молодой соединительной (грануляционной) ткани и рубцах, подвергающихся обратному развитию. Миофибробласты - особые клетки, которые по своему строению и функции занимают промежуточное положение между типичными фибробластами и клетками гладкой мышечной ткани - гладкими миоцитами. На светооптическом уровне их невозможно отличить от типичных фибробластов, однако по ультраструктурной организации они близки к гладким миоцитам, хотя, в отличие от последних, и не окружены базальной мембраной. Более половины объема их цитоплазмы занимают элементы сократительного аппарата. Их синтетический аппарат развит слабее, чем в зрелых фибробластах. Иммуноцитохимически в их цитоплазме помимо виментина выявляются актин и десмин гладкомышечного типа. Активация миофибробластов происходит при повреждении соединительной ткани. Они активно участвуют в репаративных процессах: образуют коллаген (главным образом, III типа), который заполняет и связывает поврежденные участки; сокращаясь, они стягивают края раны и уменьшают ее размеры (контракция раны - от лат. contractio - сокращение). В связи с указанной функцией миофибробласты в большом количестве обнаруживаются в молодой регенерирующей соединительной (грануляционной) ткани, рубцах, в мышечной оболочке матки при беременности. В ходе заживления раны миофибробласты с высоким содержанием актина постепенно погибают механизмом апоптоза; в рубцах они замещаются типичными фибробластами и фиброцитами. С повышенной активностью миофибробластов связывают развитие ряда заболеваний (фиброза легкого, печени, почек). Жировые клетки (адипоциты), согласно принятым представлениям, образуются из малодифференцированных (юных) фибробластов (рис. 36) путем накопления в их цитоплазме мелких липидных капель, которые сливаются между собой в одну крупную, заполняющую ее почти целиком. Жировые клетки являются нормальным компонентом рыхлой волокнистой соединительной ткани и в небольшом количестве встречаются в ней повсеместно, располагаясь по отдельности или в виде мелких скоплений. Ткань, в которой адипоциты являются структурно и функционально ведущими клеточными элементами, называют жировой и относят к одному из видов соединительных тканей со специальными свойствами. Макрофаги (гистиоциты) - вторые по численности (после фибробластов) клетки рыхлой волокнистой соединительной ткани. Они принадлежат к линии потомков стволовой клетки крови и непосредственно образуются из моноцитов после их миграции в соединительную ткань из просвета кровеносных сосудов. В соединительной ткани макрофаги располагаются поодиночке или группами. Эти клетки очень многочисленны в собственной пластинке слизистых оболочек, а также в серозных оболочках. Они могут пребывать в одном из двух взаимообратимых состояний: (1) покоящихся клеток, обладающих низкой функциональной активностью; (2) блуждающих клеток с высокой функциональной активностью. По мнению некоторых авторов, термин гистиоцит следует употреблять только применительно к клеткам в покое, однако в настоящее время он, как правило, используется в более обшем смысле для обозначения макрофага соединительной ткани. Функции гистиоцитов: 1) распознавание, поглощение и переваривание поврежденных, зараженных, опухолевых и погибших клеток, компонентов межклеточного вещества, а также экзогенных материалов и микроорганизмов; 2) участие в индукции иммунных реакций посредством захвата, переработки (процессинга) антигенов и представления их лимфоцитам (играют роль антиген-представляющих клеток); 3) регуляция деятельности клеток других типов (фибробластов, лимфоцитов, тучных клеток, эндотелиоцитов и др.). Морфологические признаки гистиоцитов зависят от степени их функциональной активности. В целом, вследствие наличия переходных форм популяция гистиоцитов характеризуются выраженным полиморфизмом. Покоящиеся гистиоциты трудно идентифицировать на светооптическом уровне. Они имеют вид мелких уплощенных клеток удлиненной или отростчатой формы с четкими контурами, прикрепленных к коллагеновым волокнам. Эти клетки характеризуются небольшим темным ядром и плотной цитоплазмой со слабо развитыми органеллами.
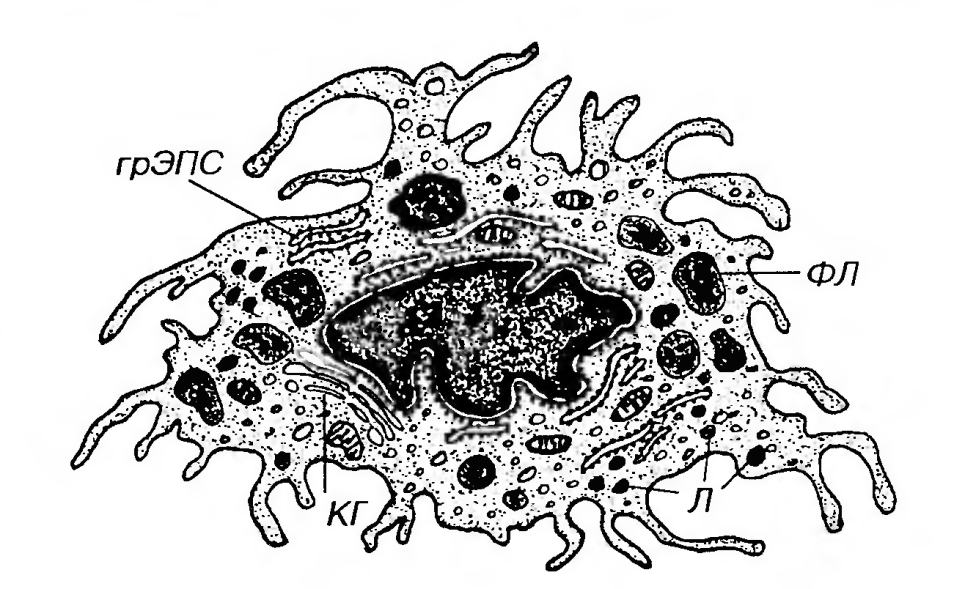
Рис. 38. Ультраструктурная организация гистиоцита. Клетка образует многочисленные цитоплазматические выросты и псевдоподии, содержит значительное число лизосом (Л) и фаголизосом (ФЛ), умеренно развитый комплекс Гольджи (КГ). Блуждающие (активные) гистиоциты обладают высокой подвижностью, изменчивой (отростчатой, реже округлой) формой с неровными, но обычно четко выявляемыми краями (рис. 35). Их ядро светлее, чем в покоящихся клетках, но темнее, чем в фибробластах; в нем может выявляться ядрышко. Цитоплазма содержит многочисленные лизосомы (рис. 38) и развитые элементы цитосклета, которые концентрируются в области псевдоподий; другие органеллы развиты умеренно. Многочисленные крупные фаголизосомы, содержащие перевариваемые продукты, в виде вакуолей хорошо видны под световым микроскопом, придавая цитоплазме гистиоцитов вспененный вид. На плазмолемме в большом количестве находятся рецепторы цитокинов, гормонов, хемоаттрактантов, а также адгезивные молекулы, которые обеспечивают контактные взаимодействия гистиоцитов с другими клетками и компонентами межклеточного вещества. Преобразования гистиоцитов в рыхлой волокнистой соединительной ткани. При активации, происходящей под действием микроорганизмов или их продуктов, а также ряда цитокинов, клетки в покое могут превращаться в блуждающие. Последние, получая стимулирующие сигналы, способны длительно находиться в состоянии высокой активности, однако в конечном итоге погибают механизмом апоптоза и фагоцитируются другими макрофагами. Под воздействием дополнительных сигналов в очаге повреждения они могут также превратиться в особые виды макрофагов - гигантские многоядерные клетки и эпителиоидные клетки. Утрачивая активность и подвижность, и прикрепляясь к коллагеновым волокнам, блуждающие клетки способны возвращаться в состояние покоя. Дендритные антиген-представляющие клетки (АПК) являются постоянными клеточными элементами рыхлой волокнистой соединительной ткани, относящимися к потомкам стволовой клетки крови. По всей видимости, они образуются непосредственно из моноцитов крови после их миграции в ткани. Не исключается полностью и возможность их развития из гематогенного предшественника, отличного от моноцитов. Установлено, что дендритные АПК в организме образуют единую систему морфологически и функционально сходных элементов. Общей функциональной особенностью дендритных АПК служит свойственная им высокая активность захвата, процессинга и представления антигенов лимфоцитам. Морфологическим признаком, характерным для этих клеток, является их отростчатая форма, наличие многочисленных ветвящихся цитоплазматических отростков, которые могут укорачиваться при перемещении клеток. Дендритные АПК, выявляемые в соединительной ткани, могут относиться к одной из двух популяций клеток: (1) АПК, специализированным на захвате антигенов только в пределах этой ткани (собственно соединительнотканным АПК), и (2) АПК, располагающимся и захватывающим антигены в эпителиях (кожи, слизистых оболочек), которые находятся в процессе миграции через соединительную ткань из эпителия в лимфатические сосуды или из кровеносных сосудов в эпителий. В ходе миграции происходят изменения ряда фенотипических свойств дендритных АПК. Тучные клетки - постоянный клеточный компонент рыхлой волокнистой соединительной ткани, осуществляющий важные регуляторные функции. Относятся к потомкам стволовой клетки крови. Тучные клетки получили свое название в связи с первоначальным ошибочным предположением о том, что их многочисленные гранулы содержат запасы питательных веществ. Этим, вероятно, объясняется и другое их название - лаброциты (от греч. labros - жадный и cytos, или kytos - клетка). Тучные клетки именуют также тканевыми базофилами, подчеркивая их сходство с базофильными гранулоцитами крови, однако это название неудачно, так как оно создает путаницу между тучными клетками и отличающимися от них базофилами крови после их миграции в соединительную ткань. Развитие тучных клеток осуществляется в тканях из предшественника, который имеет, как предполагают, костномозговое происхождение. На их дифференцировку и рост влияют ИЛ-3 (продуцируемый Т-лимфоцитами) и факторы клеточного микроокружения (фибробласты, эпителиальные клетки и их продукты). В отличие от базофилов, которые после миграции в ткани живут недолго (от нескольких часов до нескольких суток), тучные клетки, по-видимому, обладают сравнительно большой продолжительностью жизни (от нескольких недель до нескольких месяцев). В течение этого периода под действием соответствующих стимулов тучные клетки, очевидно, способны делиться. Функции тучных клеток в целом сходны с функциями базофилов, находящихся в тканях. К ним относятся: 1) Гомеостатическая, которая осуществляется в физиологических условиях путем медленного выделения небольших количеств биологически активных веществ, способных влиять на различные тканевые функции - в первую очередь, на проницаемость и тонус сосудов, и поддержание баланса жидкостей в тканях. 2) Защитная и регуляторная, которая обеспечивается путем локального выделения медиаторов воспаления и хемотаксических факторов, обеспечивающих (а) мобилизацию эозинофилов и различных эффекторных клеток, участвующих в так называемых реакциях поздней фазы; (б) воздействие на рост и созревание соединительной ткани в зоне воспаления. 3) Участие в развитии аллергических реакций вследствие наличия высокоаффинных рецепторов к иммуноглобулинам класса Е (IgE) на их плазмолемме и функциональной связи этих рецепторов с секреторным механизмом. Распределение тучных клеток в организме. Тучные клетки располагаются преимущественно около мелких сосудов - периваскулярно, что, вероятно, связано с их регуляторной функцией и влиянием на проницаемость сосудов. Распределение тучных клеток в организме неравномерно - соединительная ткань различных органов содержит неодинаковое их количество. Этими клетками особенно богата дерма. Они также очень многочисленны в собственной пластинке слизистых оболочек пищеварительного тракта, дыхательной, выделительной и половых систем, в строме молочной железы и тимуса. В среднем, в рыхлой волокнистой соединительной ткани их относительное содержание составляет 10% от общего числа клеток. Проявлением регуляторной функции тучных клеток служит нарастание их количества в строме различных органов, функциональная активность которых повышается, например, в щитовидной железе при ее гиперфункции, в лактирующей молочной железе, в матке при беременности и в течение менструального цикла и т.п. Оно увеличено также вблизи и внутри очагов хронического воспаления, в опухолях и по периферии заживающих ран. Механизмами локального нарастания содержания тучных клеток могут служить их миграция, обусловленная хемоаттрактантами, усиленная дифференцировка из местных предшественников и, возможно, митотическое деление. В тканях тучные клетки устанавливают многочисленные адгезивные контакты с фибробластами, эндотелиалъными клетками мелких сосудов, коллагеновыми и нервными волокнами, молекулами фибронектина, ламинина и другими компонентами межклеточного вещества. Эти взаимодействия оказывают регуляторные влияния как на состояние самих тучных клеток (способствует их дифференцировке из предшественников, облегчают их миграцию, распластывание, секреторную реакцию), так и на клетки других типов. Строение тучных клеток. Тучные клетки имеют удлиненную или округлую форму, неровную поверхность с многочисленными тонкими отростками и выростами. Они в 1,5-2 раза крупнее базофилов (диаметр 20-30 мкм). Ядро тучных клеток - сравнительно небольшое, несегментированное, овальное или округлое, с умеренным содержанием гетерохроматина. На светооптическом уровне оно часто прослеживается с трудом, так как маскируется гранулами, содержащимися в цитоплазме. Цитоплазма тучных клеток содержит умеренно развитые органеллы, элементы цитоскелета, липидные капли и гранулы (рис. 39).
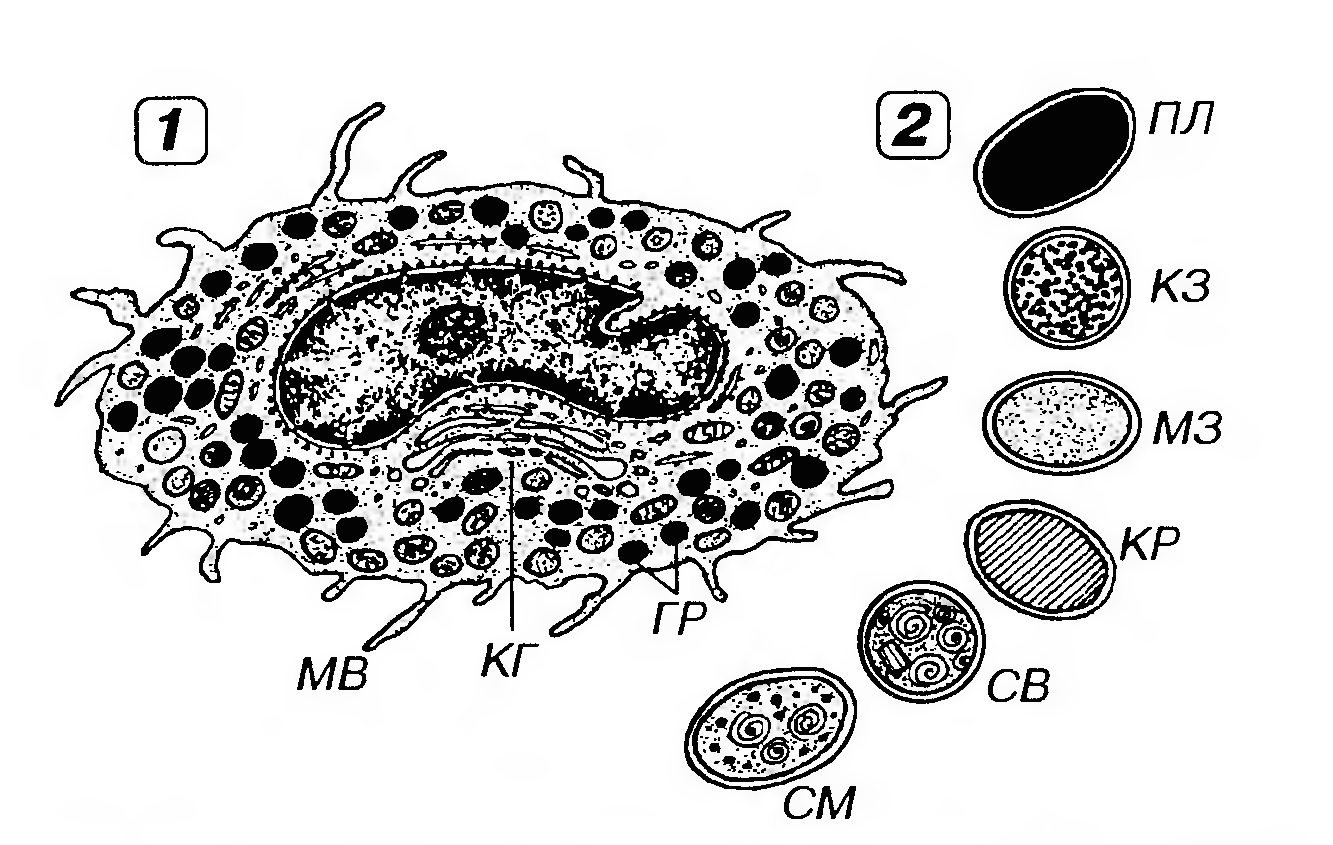
Рис. 39. Ультраструктурная организаций тучной клетки (1) и морфологическая вариабельность содержимого ее гранул (2). MB - микроворсинки, КГ - комплекс Гольджи, ГР - гранулы, с плотным (ПЛ), крупнозернистым (КЗ), мелкозернистым (МЗ) гомогенным содержимым, с кристаллоидной структурой (КР), с матриксом, содержащим структуры в виде "пергаментных свитков" (СВ), смешанного строения (CM). Гранулы тучных клеток сходны по строению и составу содержимого с гранулами базофилов, но не идентичны им. Они также окрашиваются метахроматически, но они мельче, чем в базофилах, более многочисленны и обладают более варабельной формой и ультраструктурой (даже в составе одной клетки). Встречаются гранулы с плотным, крупно- или мелкозернистым гомогенным содержимым, с кристаллоидной структурой, с матриксом умеренной плотности, в который погружены более плотные структуры (иногда в форме "пергаментных свитков"). Последний вид гранул особенно характерен для тучных клеток слизистых оболочек. Нередко обнаруживаются гранулы смешанного строения (рис. 39). Содержимое гранул тучных клеток: гепарин, гистамин, дофамин, хемотаксические факторы эозинофилов и нейтрофилов, хондроитинсульфаты, гиалуроновая кислота, гликопротеины и фосфолипиды. В составе основных белков гранул имеются нейтральные протеазы, кислые гидролазы, катепсин G. Функциональная морфология тучных клеток в физиологических условиях. Феномен медленной дегрануляции тучных клеток человека (длящейся сутками), как и аналогичная реакция базофилов, установлен лишь в последние годы. Ее структурным механизмом служит микровезикулярный транспорт содержимого специфических гранул к плазмолемме. Малые дозы биологически активных веществ, выделяющиеся при медленной секреции, обусловливают локальные физиологические регуляторные реакции, направленные на поддержание гомеостаза (преимущественно на изменения тонуса и проницаемости сосудов, а, следовательно, активности трофики тканей и водно-солевого баланса). Структурно-функциональные различия тучных клеток. Популяция тучных клеток образована элементами, которые обладают неодинаковыми морфофункциональными свойствами и могут качественно и количественно различаться даже в пределах одного органа. Высказывают предположение о том, что отдельные субпопуляции тучных клеток выполняют в организме неодинаковые функции. Типы тучных клеток различают на основании особенностей окраски и содержания медиаторов в их гранулах, ультраструктуры, количества рецепторов на плазмолемме (и, следовательно, чувствительности к действию различных угнетающих и стимулирующих факторов), активности ряда ферментов. Описаны клетки двух основных типов: (1) тучные клетки соединительной ткани (находятся преимущественно в составе дермы и стромы различных органов); (2) тучные клетки слизистых оболочек (преобладают в собственной пластинке слизистых оболочек). Дифференцировка предшественников тучных клеток в тот или иной тип зрелых клеток определяется, как предполагают, факторами микроокружения и влиянием цитокинов. Участие тучных клеток в развитии аллергических реакций, как и базофильньгх гранулоцитов, включает: (1) связывание IgE с высокоаффинными рецепторами на их плазмолемме; (2) взаимодействие мембранного IgE с аллергеном; (3) активацию и дегрануляцию тучных клеток с выделением содержащихся в их гранулах веществ и продукцией ряда новых. Дегрануляция может опосредоваться также рецепторами комплемента или вызываться белками нейтрофилов, протеиназами, нейропептидами (вещество Р, соматостатин), лимфокинами. Активация тучных клеток индуцирует синтез и выделение ими эйкозаноидов - производных ненасыщенных жирных кислот (простагландинов, тромбоксана, простациклина и лейкотриенов), играющих важную роль в сосудистых реакциях, сокращении гладких мышц внутренних органов и привлечении нейтрофилов. Продуцируемый ими ФАТ (фактор, активирующий тромбоциты), вызывает гиперреактивность бронхов, усиливает сосудистую проницаемость, отек и инфильтрацию ткани тучными клетками и эозинофилами. Выработка цитокинов тучными клетками. Тучные клетки продуцируют разнообразные мультифункциональные цитокины (ФНОα, ИЛ-1, -2, -3, -4, -5, -6, ГМ-КСФ и др.), которые накапливаются в их гранулах или вновь синтезируются при активации. Эти вещества оказывают действие на многие типы клеток, участвующих в различных процессах, в частности, в так называемых реакциях поздней фазы - длительной иммунной стимуляции, развивающейся спустя несколько часов после контакта с аллергеном. Анафилактическая дегрануляция тучных клеток человека протекает в течение нескольких минут. Она начинается с набухания гранул, содержимое которых частично растворяется и становится менее плотными, в дальнейшем гранулы сливаются в извитые цепочки. Последние превращаются во внутрицитоплазматические каналы, содержащие материал гранул. Мембрана каналов (или реже отдельных набухших гранул) сливается с плазмолеммой, обеспечивая выделение их содержимого за пределы клетки. Множественные устья каналов расширяются, что сопровождается образованием многочисленных складок и выпячиваний плазмолеммы. Восстановление исходных морфологических особенностей тучных клеток после дегрануляции (регрануляция) занимает более 24-48 ч. Результатом анафилактической дегрануляции тучных клеток, как и базофилов, служат разнообразные реакции, связанные со спазмом гладких мышц, расширением сосудов, повышением их проницаемости, повреждением тканей (например, эпителия бронхов, кишки). Выделение различных ферментов (протеаз, карбоксипептидаз и др.) обусловливает переваривание компонентов межклеточного вещества с образованием веществ, обладающих хемотаксическим действием на гранулоциты, макрофаги и фибробласты. Клинические проявления массивной дегрануляции тучных клеток зависят от распространенности и преимущественной тканевой и органной локализации этой реакции. Они включают бронхоспазм, острый ринит, отеки, кожный зуд, понос, падение кровяного давления вплоть до анафилактического шока и смерти. Участие тучных клеток в реакциях поздней фазы (длительной иммунной стимуляции). Разнообразные биологически активные вещества, выделенные тучными клетками, привлекают базофилы, эозинофилы, нейтрофилы, макрофаги, а также другие клетки и облегчают их миграцию из кровеносных сосудов в ткани, усиливая их адгезию к эндотелию. Выселившиеся клетки секретируют ряд собственных медиаторов, которые могут привлекать новые клетки, поддерживая или усугубляя повреждение тканей. Вместе с тем, некоторые из продуцируемых тучными клетками веществ способствуют течению репаративных процессов, в частности, стимулируют выработку межклеточного вещества фибробластами и ангиогенез. Вещества, угнетающие дегрануляцию тучных клеток, (с различным механизмом фармакологического действия) нашли широкое клиническое применение в качестве средств профилактики и лечения аллергических заболеваний. Плазматические клетки (плазмоциты) и их предшественники - В-лимфоциты, находящиеся на различных этапах преобразования в плазмоциты - в небольших количествах постоянно содержатся в различных участках рыхлой волокнистой соединительной ткани (рис. 35). Они особенно многочисленны в соединительной ткани серозных оболочек, собственной пластинки различных слизистых оболочек, а также вокруг концевых отделов и выводных протоков экзокринных желез. Эти клетки имеют мелкие размеры, располагаются поодиночке или группами и обладают высокой синтетической и секреторной активностью, вырабатывая и выделяя антитела (иммуноглобулины) и обеспечивая тем самым гуморальный иммунитет. Лейкоциты (гранулоциты и агранулоциты) являются нормальными клеточными компонентами рыхлой волокнистой соединительной ткани, в (или через) которую они мигрируют для выполнения своих функций после выхода из кровеносного русла. Лимфоциты, в отличие от других видов лейкоцитов, способны из соединительной ткани через оттекающую лимфу вновь попадать в кровь (осуществлять рециркуляцию). Содержание лейкоцитов в рыхлой волокнистой соединительной ткани в норме незначительно. Выделяя цитокины, эти клетки могут оказывать регуляторное влияние друг на друга, на остальные виды клеток соединительной ткани и на клетки соседних тканей. Локальное увеличение числа лейкоцитов в рыхлой волокнистой соединительной ткани, образующих в большей или меньшей степени очерченные скопления, выявляется при воспалении. В частности, при остром воспалении в таких скоплениях (инфильтратах) преобладают нейтрофильные гранулоциты, при хроническом - обнаруживаются преимущественно лимфоциты, плазматические клетки, моноциты и образующиеся из них макрофаги. Пигментные клетки человека имеют нейральное происхождение и являются потомками клеток, выселившихся в эмбриональном периоде из нервного гребня. Цитоплазма этих клеток содержит пигменты меланины (от греч. melanos - черный). Цвет пигментов варьирует от коричнево-черного (эумеланины) до желто-коричневого (феомеланины). Пигментные клетки имеют отростчатую форму и подразделяются на два вида - меланоциты, которые вырабатывают пигмент, и меланофоры, способные лишь накапливать его в цитоплазме. Пигментные клетки входят в состав рыхлой волокнистой соединительной ткани (рис. 35), хотя у человека и других млекопитающих они встречаются в ней сравнительно редко. Повышенное содержание пигментных клеток характерно для соединительнотканной части кожи (дермы) некоторых участков тела (мошонки, сосков, перианальной области). Содержание пигментных клеток в соединительной ткани дермы увеличено в области родимых пятен (невусов). Численное преобладание пигментных клеток над другими клеточными элементами соединительной ткани характерно для радужки и сосудистой оболочки глаза, где им принадлежит и функционально ведущая роль. Такую ткань называют пигментной и относят к одному из видов соединительных тканей со специальными свойствами.
|
||
|
|
Последнее изменение этой страницы: 2021-02-07; просмотров: 1070; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.019 с.) |