Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Характеристика минорных классов веществ вторичного метаболизма растений. Распространение и основные представители.Содержание книги
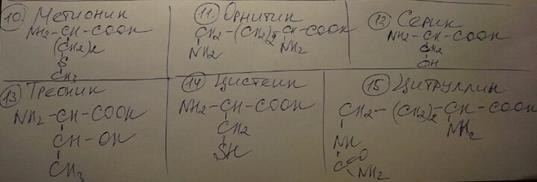
Поиск на нашем сайте Гликозиды — широко распространенные в природе вещества, в молекулах которых остатки сахара связаны с молекулой вещества неуглеводной природы — агликоном. Агликонами могут быть самые различные соединения (спирты, кислоты, фенольные соединения, амины и др.), поэтому гликозиды, как правило, не расcматриваются в качестве отдельного класса вторичных метаболитов. Так, например, гликозиды, содер-жащие в качестве агликона фенольные соединения (антоцианы, халконы), принято относить к фенолам. Гликозиды, содержащие в качестве агликонов соединения стероидной природы (сердечные гликозиды и сапонины), относят к изопреноидам (тритерпеноидам). Цианогенные гликозиды являются β-гликозидами 2-гидрок- синитрилов (цианогидринов). Они образуются из аминокислот и содержат в своем составе синильную кислоту. В качестве углеводного фрагмента у цианогенных гликозидов, как правило, выступает D-глюкоза. Цианогенные гликозиды присутствуют, как правило, в вакуолях клетки, а β-гликозидаза, отщепляющая углеводный фрагмент молекулы, — в цитозоле. Следует отметить, что для β-гликозидаз характерна избирательность по отношению к различным циа- ногенным гликозидам. В частности, гликозидазы, достаточно активно расщепляющие ароматические цианогенные гликозиды, гораздо менее активны по отношению к гликозидам с алифатическимиагликонами (линамарин, лотаустралин). Тиольные гликозиды содержат в своем составе тиоцианат-ные, изотиоцианатные, сульфо- и неорганические агликоны [см.: 6, с. 125]. При гидролизном распаде образуют соответствующие сахара и меркаптаны. Тиольные гликозиды широко представлены в растениях семейства крестоцветных. Они находятся в виде солей с щелочными металлами, чаще всего с калием. Обладают сильным антимикробным действием и обусловливают острый или жгучий вкус горчицы, хрена, редьки. В растении тиольные гликозиды, как и цианогенные гликозиды, пространственно отделены от гидролизующих их ферментов. Растительные амины. Амины можно рассматривать как производные аммония. Различают четыре основные группы аминов: первичные, вторичные, третичные и четвертичные. Следует отметить, что достаточно сложно четко отделить растительные амины от других классов вторичных соединений, прежде всего алкалоидов. Четких критериев отличия биогенных аминов от протоалкалоидов нет. В группу биогенных аминов обычно не включают соединения с карбоксильными группами, а также пурины и пиримидины. Многие из аминов структурно представляют собой декарбоксилированные аминокислоты, причем как протеиногенные, так и непротеиногенные. Растительные амины обычно разделяют на моноамины (с одной аминогруппой), диамины (с двумя аминогруппами) и полиамины (более двух аминогрупп). Амины — ядовитые вещества, которые могут вызывать отравления как животных, так и растений. Они в малых количествах обнаружены в грибах (рожки спорыньи, мухоморы, дрожжи) и в растениях (дурман, белена, омела, соя). Амины, образующиеся при декарбоксилировании аминокислот, обычно в растениях не накапливаются, а претерпевают различные превращения и вовлекаются в обмен веществ. Непротеиногенные аминокислоты Под термином «непротеиногенные» (небелковые) аминокислоты подразумевают природные аминокислоты, их амиды, ими-нокислоты, которые, как правило, не входят в состав белков. Большинство непротеиногенных аминокислот в высших растениях находятся в свободном состоянии или в конденсированном виде с другими низкомолекулярными соединениями (глутаминовой, щавелевой, уксусной кислотами). Некоторые из непротеино- генных аминокислот, например, гидроксипролин или гомосерин, встречаются в составе растительных белков. Однако подобные случаи достаточно редки, при этом белки с такими аминокисло-тами выполняют, как правило, специфические функции (например гидроксипролин-обогащенные белки в клеточных стенках). К тому же образование непротеиногенных аминокислот в подобных белках происходит чаще всего путем посттрансляционной модификации протеиногенных аминокислот (например, гидроксипролин образуется путем гидроксилированияпролина). При этом наиболее обычными вариациями являются: – удлинение или сокращение углеродной цепи (добавление или удаление СН2 - или СН3 -фрагментов); – гидрирование и дегидрирование; – гидроксилирование; – аминирование. Фитохимическую классификацию непротеиногенных аминокислот обычно связывают со структурой «исходных» протеиногенных аминокислот. При этом различают: – нейтральные алифатические аминокислоты; – серосодержащие аминокислоты; – ароматические аминокислоты; – гетероциклические аминокислоты; – дикарбоновые аминокислоты и амиды; – щелочные (основные) аминокислоты; – иминокислоты. Другая классификация непротеиногенных аминокислот осно-вана на характере сходства с протеиногенными. Выделяют три основные группы. - Сходство по изомерии; - Сходство по гомологии; - Сходство по аналогии. Непротеиногенные аминокислоты выполняют в растениях ряд важных функций: – некоторые из них участвуют в образовании протеиногенных. – непротеиногенные аминокислоты могут служить запа-сной формой азота и серы. – некоторые непротеиногенные аминокислоты (например, производные аспарагиновой и глутаминовой кислот и их амидов) являются транспортной формой азота. – непротеиногенные аминокислоты могут выполнять разнообразные защитные функции. Беталаины являются еще одной группой (помимо антоци-анов) водорастворимых пигментов высших растений. По своей химической структуре они представляют собой алкалоиды. Это единственная группа алкалоидов, имеющих яркую окраску. Беталаины встречаются в виде гликозидов, находятся в вакуо- лях и разделяются на бетацианины и бетаксантины. Бетацианины являются красно-фиолетовыми водораство-римыми пигментами растений порядка Caryophyllalles. Бетаксантины являются желтыми водорастворимыми пигмен-тами растений порядка Caryophyllalles. Подобно повсеместно распространенным антоцианам, беталаины находятся в вакуолях клеток цветков, плодов и листьев. Необычные липиды. Многие липиды растительного происхождения имеют признаки вторичных метаболитов: они являются низкомолекулярными соединениями, встречаются лишь вопределенных таксономических группах растений, многие из них обладают биологической активностью. Жирные кислоты. Необычные жирные кислоты являются типичными вторичными метаболитами: они встречаются лишь в растениях определенных таксономических групп и не являются компонентами мембран клеток. Необычные жирные кислоты отличаются от «обычных» длиной углеродной цепи, необычным расположением и количеством двойных связей, наличием дополнительных функциональных групп и циклов. В углеродных алифатических цепях могут находиться заместители различной химической природы, такие как метильные, гидроксильные, карбонильные группы, оксо- и эпоксигруппы, циклопропановые или циклопентановые группировки. В эту же группу включены жирные кислоты с тройными связями, конъюгированными двойными связями, с разветвленной углеродной цепочкой, с двойными связями в транс-форме. Чаще всего необычные жирные кислоты обнаруживаются в масле семян. Примером «коротких» насыщенных жирных кислот может служить каприевая кислота, имеющая 12 углеродных атомов и обнаруженная в представителях семейств Ulmaceae, Lauraceae, Lythraceae. Длинноцепочечная насыщенная жирная кислота с 24 атомами углерода — лигноцериновая — найдена в растениях семейств Leguminosae и Sapindaceae. Производные ацетилена У многих видов высших растений при изучении липидного спектра были обнаружены соединения с одной или несколькими тройными связями. Такие соединения получили название ацетиленовых производных. Изановая кислота, имеющая две тройные и одну двойную связь, найдена в представителях семейства Olacaceae. Ацетиленовые производные распространены очень широко: они обнаружены у растений, животных и микроорганизмов. Их можно разделить на длинноцепочечные моноацетилены и полиацетилены. Полиацетиленовые производные более разнообразны в структурном плане, поэтому получили более широкое распространение. Воска. Эпидермис различных органов высших растений, как правило, покрыт кутикулой, на которой обычно имеется восковой налет различной толщины. Этот восковой слой носит название кутику-лярного воска. Растительные воска имеют сложный состав. Кути-кулярный воск представляет собой многокомпонентную смесь, которая состоит из относительно простых углеводородов (прежде всего, алканов), восковых эфиров, а также жирных кислот, спиртов и кетонов. 31. Непротеиногенные аминокислоты. Пути их образования и функции. Это аминокислоты, их амиды, иминокислоты, которые в норме не входят в состав белков. В растениях и микроорганизмах, в частности, они образуются вовремя повышенной потребности в азоте, например, при образовании почек или прорастании семян, или же запасаются в виде растворимых веществ. Многие аминокислоты, образовавшиеся при обмене веществ низших организмов, имеют свойства антибиотиков. Они действуют как аминокислоты - антагонисты, т.е. являются конкурентными ингибиторами при обмене веществ, задерживая определенные ступени биосинтеза аминокислот или способствуя образованию ложных последовательностей при биосинтезе белков. Больш-во Н.АК (непротеиног аминок-ты) в высших растениях находятся в свободном состоянии или в конденсированном виде с другими низкомолекулярными соединениями (глутаминовой, щавелевой, уксусной кислотами). Некоторые из Н.АК (гидроксипролин или гомосерин) встречаются в составе растительных белков. Например, гидроксипролин обогащае белки в клеточных стенках. Образование Н.АК в белках в этом случае происходит путем посттрансляционной модификации протеиногенных АК. Известно более 400 непротеиногенных аминокислот. Многие из них – модифицированные белковые аминокислоты. Так, аланину соответствуют свыше 30 производных, различающихся заместителями водородного атома ме-тильной группы. Заместителем может быть аминогруппа, как, например, у 1,2-диаминопропановой кислоты H2NCH2CH(NH2)COOH, которая встречается в растениях семейства мимозовых; может образоваться циклопропановое кольцо, как у найденной в различных фруктах аминокислоты гипоглицина и 1-аминоциклопропакарбоновой кислоты. Осн.пути синтеза непротеиногенных: 1. Удлинение или сокращение цепи (добавление/удаление СН2-, СН3-фрагмента). 2. Гидрирование, дегидрирование. 3. Гидроксилирование. 4. Аминирование Существует фотохимическая классификация Н.АК, связанная со структурой исходных протеиногенных АК.: 1. Нейтральные алифатические АК. 2. Серосодержащие АК. 3. Ароматические АК. 4. Гетероциклические АК. 5. Дикарбоновые АК и амиды. 6. Щелочные (основыные) АК. 7. Иминокислоты. Классификация небелковых аминок-т по сходству с белковыми: 1. Сходство по изомерии (альфа- аланин (протеин.)и бета-аланин (Н. АК)). Бета-аланин сод-ся в пуле свободных аминокислот, входит в состав пантотеновой кислоты, коэнзима А. 2. По гомологии (цистеин-гомоцитеин, аргинин-гомоаргинин, серин-гомосерин). 3. По аналогии (аналогия по замещению - Цистеин и S-метилцистеин (он распространен в растениях и может выступать донором метильных групп в реакциях метилирования). Либо по сходству молекул – канаванин и аргинин. Функции непротеиногенных аминокислот в растениях: 1. Транспортная форма азота по растению. Производные асп и глу и их амидов. 2. Основная запасная форма азота и серы в семенах. Орнитин, цитруллин, гомоаргинин накапливаются в семенах в качестве резерва азота. При прорастании используются для образования необходимых проростку аминокислот. Источник запасной серы - S-метилцистеин. 3. Быстрая и эффективная регуляция количества доступного NH4+ для растения. Непротеиногенные аминокислоты легко подвержены метаболизму и в случае необходимости снабжают растение ионами аммония для синтеза белков. 4. Некоторые непротеиногенные аминок-ты участвуют в образовании протеиногенных. Например, из гомосерина образуется треонин и метионин. 5. Защитная функция. Например, орнитин и цитруллин участвуют в обезвреживании аммиака в орнитиновом цикле. Принимают участие в синтезе этилена. Метионин – аминоциклопропановая кислота (Н.АК) – явл-ся транспортоной формой этилена – этилен. 6. Непротеиногенные аминокислоты могут быть даже токсичны для организма другого вида, они ведут себя как обычные чужеродные вещества (ксенобиотики). Например, гипоглицин, канаванин, дьенколевая кислота и β-цианоаланин, выделенные из растений, ядовиты для человека. Токсичность обусловлена тем, что они включаются в состав белков вместо нормальных белковых АК и нарушают функционирование таких протеинов. Например, токсичность канаванина связана с тем, что у животных аминоацит-тРНК-синтазы, осуществляющие перенос аргинина на тРНК, не могут отличить канаванин от аргинина и включают его в цепь. Формулы, встреченных в тексте АК (по алфавиту).
|
||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 418; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.01 с.) |