Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Биологические функции липидовСодержание книги
Поиск на нашем сайте 1. Макроэргические вещества. Липиды —наиболее важный из всех питательных веществ источник энергии. В основном жир содержится в клетках в виде жировых капель, которые служат метаболическим «топливом». Липиды окисляются в митохондриях до воды и диоксида углерода с образованием большого количества АТФ. 2. Структурные блоки. Ряд липидов принимает участие в образовании клеточных мембран.Типичными мембранными липидами являются фосфолипиды, гликолипиды и холестерин. Стерины у эукариот стабилизируют внутриклеточные мембраны. 3. Изолирующий материал. Жировые отложения животных в подкожной ткани и вокруг различных органов обладают высокими теплоизолирующими свойствами. Как основной компонент клеточных мембран липиды изолируют клетку от окружающей среды и за счет гидрофобных свойств обеспечивают формирование мембранных потенциалов. 4. Прочие функции липидов. Стероиды выполняют сигнальные функции, служат в качестве гормонов, медиаторов и вторичных переносчиков (мессенджеров). Отдельные липиды выполняют роль «якоря», удерживающего на мембране белки и другие соединения. Некоторые липиды являются кофакторами, принимающими участие в ферментативных реакциях, например, в свертывании крови или в трансмембранном переносе электронов. Светочувствительный каротиноид ретиналь играет центральную роль в процессе зрительного восприятия. Функции в растениях: энергетическая, защитная, регуляторная, структурная. Регуляторная: фитогормоны; жирорастворимые витамины. Защитная: воска защищают растения от высыхания, воздействий внешней среды и микроорганизмов (образуют тонкий слой на поверхности листьев, стеблей и плодов растений). Фотосинтетические пигменты. Изопреноиды принимают участие в переносе электронов в ф/с и дыхательной электронтранспортных цепях (пренильные бок.цепи пластохинона и убихинона). Структура и свойства. Полярные гидрофильные головки и неполярные гидрофобные хвосты. Двойственная природа этих липидов обусловливает их ключевую роль в организации биологических мембран. Атомы углерода в молекулах высших карбоновых кислот могут быть соединены друг с другом как простыми, так и двойными связями. Из предельных (насыщенных) жирных кислот наиболее часто в состав жиров входят пальмитиновая, стеариновая, арахиновая; из непредельных (ненасыщенных) — олеиновая и линолевая. Степень ненасыщенности и длина цепей высших карбоновых кислот (т.е. число атомов углерода) определяют физические свойства того или иного жира. Жиры с короткими и непредельными кислотными цепями имеют низкую температуру плавления. При комнатной температуре это жидкости(масла). И наоборот, жиры с длинными и насыщенными цепями жирных кислот при комнатной температуре твердые. Вот почему при гидрировании (насыщении кислотных цепей атомами водорода по двойным связям) жидкое арахисовое масло, например, становится мазеобразным, а подсолнечное масло превращается в твердый маргарин. В фосфолипидах одна из крайних цепей высших карбоновых кислот триацилглицерола замещена на группу, содержащую фосфат. Гликолипиды образуются в результате соединения липидов с углеводами.
(гликосфинголипид) (фосфолипид)
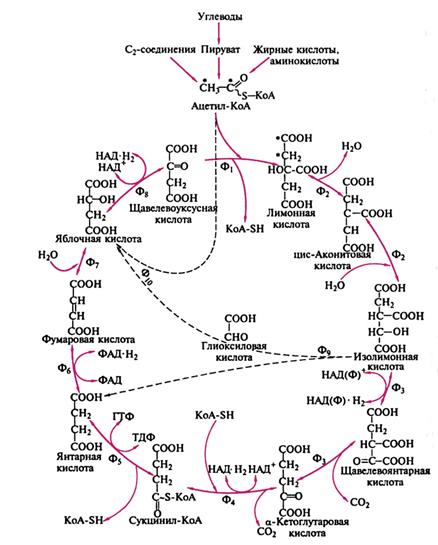
Глиоксилатный шунт. Глиоксилатный цикл - циклический ферментативный процесс, в котором происходит превращение ацетил-KoA в различные вещества, необходимые для синтеза всех важных клеточных компонентов, через стадию образования янтарной и глиоксиловой кислот. Является модификацией цикла Кребса и локализован не в митохондриях, а в мембраносвязанных органеллах - глиоксисомах. В этих органеллах образуется изолимонная кислота, как и в цикле Кребса. Затем она под действием изоцитратлиазы распадается на глиоксиловую и янтарную кислоты. Глиоксиловая кислота реагирует со второй молекулой ацетилкоэнзима А с образованием яблочной кислоты, которая затем окисляется до щавелевоуксусной кислоты. Янтарная кислота выходит из глиоксисомы и превращается в щавелевоуксусную кислоту. В ходе глиоксилатного цикла утилизируются две молекулы ацетилкоэнзима А, образовавшегося при распаде запасных жиров, и образуется одна молекула НАДН.У растений ферменты глиоксилатного цикла находятся в глиоксисомах, представляющих собой специализированные пероксисомы (названы по протекающему в них циклу). Ферменты, одновременно участвующие и в цикле трикарбоновых кислот, и в глиоксилатном цикле, имеют по два изофермента, один из которых локализуется в митохондриях, а другой — в глиоксисомах. Глиоксисомы не присутствуют постоянно во всех растительных тканях. Они образуются в богатых липидами семенах при их прорастании, ещё до того, как развивающееся растение сможет само образовывать глюкозу в ходе фотосинтеза. Кроме ферментов глиоксилатного цикла, в глиоксисомах находятся все ферменты, необходимые для деградации жирных кислот, запасённых к семенах. Ацетил-СоА, образующийся при β-окислении жирных кислот, превращается в сукцинат в ходе глиоксилатного цикла, и сукцинат транспортируется в митохондрии, где он вовлекается в цикл трикарбоновых кислот и превращается в малат. Цитозольный изофермент малатдегидрогеназы окисляет малат до оксалоацетата, который в свою очередь может превратиться в фосфоенолпируват. Последний вовлекается в глюконеогенез и даёт в конечном итоге глюкозу. Таким образом, прорастающее семя может переводить углерод, запасённый в липидах, в глюкозу.
Ф1 - цитратсинтаза (конденсирующий фермент); Ф2 - аконитаза; Ф3 - изоцитратдегидрогеназа; Ф4 - альфа-кетоглутаратдегидрогеназа; Ф5 - сукцилтиокиназа; Ф6 - сукцинатдегидрогеназа; Ф7 - фумараза; Ф8 - малатдегидрогеназа; Ф9 - изоцитратлиаза; Ф10 - малатсинтетаза. Включение углеродных атомов ацетильного остатка в молекулу лимонной кислоты помечено звездочками. Пунктирными линиями изображены реакции глиоксилатного шунта.
|
||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 331; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.147 (0.007 с.) |