
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лекція 3. Будова еукаріотичної клітини
Будова і функції плазмолеми.. Цитоплазма. Склад цитоплазми. Гіалоплазма, органели і включення. Класифікація органел. Органели загального та спеціального призначення, мембрані та немембранні органели, мікро- і ультрамікроскопічні органели. Будова та функції ендоплазматичної сітки, мітохондрій і комплексу Гольджі.
Еукаріотна клітина складається з плазмолеми (оболонки), цитоплазми та ядра (рис. 3). Плазмолема має товщину 10 нм. Її можна розглянути тільки за допомогою електронного мікроскопа. Вона обмежує клітину і складається з трьох шарів: зовнішнього, середнього й внутрішнього. Зовнішній шар — глікокалікс, або надмембранний комплекс, середній — клітинна мембрана і внутрішній — підмембранний комплекс (рис. 4). Будову клітинної мембрани описано вище. Глікокалікс утворений переважно вуглеводами — олігоцукридами, які утворюють з білками й ліпідами клітинної мембрани складні сполуки — відповідно глікопротеїди та гліколіпіди.
Рис. 3. Загальний план будови клітини (схема) Вуглеводні кінці молекул цих сполук утворюють довгі розгалужені ланцюги. Глікокалікс виконує рецепторну функцію плазмолеми. За його участю здійснюється взаєморозпізнавання клітин та їх взаємодія з навколишнім середовищем. У глікокаліксі епітеліоцитів кишок сконцентровані ферменти пристінного травлення. Підмембранний комплекс представлений мікрофіламентами й мікротрубочками, які розміщені в периферичній частині клітини і є частиною цитоскелета. Структури цього комплексу мають здатність взаємно переміщуватись, що спричинює переміщення плазмолеми, частин клітини й клітини. Підмембранний комплекс бере також участь у рецепції й трансмембранному транспорті. Плазмолема виконує численні функції. Основними з них є розмежувальна, транспортна, рецепторна, рухова та формування клітинних контактів. Розмежувальна і транспортна функції є взаємопротилежними і взаємодоповнювальними. Завдяки розмежуванню з навколишнім середовищем клітина зберігає свою індивідуальність, завдяки транспорту речовин здійснюються метаболічні процеси як у самій клітині, так і між клітиною та середовищем, що її оточує. У результаті цього забезпечується сталість внутрішнього середовища клітини. Вода і розчинені в ній гази, окремі йони та дрібні молекули органічних речовин транспортуються в клiтину i з неї шляхом дифузії (пасивний транспорт). Більшість йонів і невеликих молекул неорганічних та органічних речовин (солі, цукри, амінокислоти) транспортуються проти градієнта концентрації за участю особливих ферментів — пермеаз (активний транспорт супроводжується витрачанням енергії).
Великі молекули органічних речовин та їх агрегати проникають у клітину в результаті процесу ендоцитозу, який поділяють на фагоцитоз і піноцитоз. Фагоцитоз — процес захоплення й поглинання клітиною великих часточок (бактерії, частинки пилу, фрагменти інших клітин). У процесі піноцитозу клітиною захоплюються й поглинаються макромолекулярні сполуки у вигляді розчинів. Часточки, що поглинаються, обволікаються плазмолемою і у вигляді мішечка втягуються всередину цитоплазми. Частини плазмолеми, які оточують поглинуті субстанції, втрачають зв’язок з плазмолемою клітини, внаслідок чого утворюються фагосоми й піноцитозні пухирці. Вони з’єднуються з лізосомами, під дією ферментів яких поглинуті часточки перетравлюються і засвоюються клітиною. Виведення клітиною речовин за межі цитоплазми називають екзоцитозом. Він має низку різновидів: секреція — виведення синтезованих продуктів; екскреція — виведення шкідливих або токсичних речовин; рекреція — виведення речовин, які не змінюють своєї хімічної структури в процесах метаболізму (вода, солі); клазматоз — видалення окремих структурних компонентів клітин. Рецепторну функцію плазмолеми виконують глікокалікс і клітинна мембрана. Рух плазмолеми забезпечують елементи підмембранного комплексу, які здатні змінювати своє положення. Ферменти, що містяться в глікокаліксі плазмолеми ентероцитів, зумовлюють функцію пристінного травлення. Плазмолема бере участь у формуванні міжклітинних контактів. Вони добре виражені в клітинах, які прилягають одна до одної (епітеліальні та ендотеліальні клітини). Залежно від особливостей будови, зумовлених їх функціями, міжклітинні контакти поділяють на: прості — між плазмолемами є щілина завширшки 15 – 20 нм, щільні — плазмолеми клітин максимально зближені, десмосоми, або плями злипання, — фібрилярні структури цитоплазми контактуючих клітин утворюють пластинки прикріплення, зубчасті — плазмолеми утворюють вирости, які заходять у розміщені між ними заглибини, щілинні — між клітинами є щілина завширшки 2 – 3 нм, а в їх плазмолемі знаходяться особливі канали, через які проникають йони та дрібні молекули.
Цитоплазма клітини складається з гіалоплазми, органел і включень. Гіалоплазма — це найбільш рідка частина цитоплазми. Вона становить близько 50 % загального об’єму цитоплазми. До складу гіалоплазми входить вода з розчиненими в ній мінеральними та органічними речовинами. Високомолекулярні речовини гіалоплазми у вигляді ниток утворюють ніжну суцільну тривимірну сітку, яка формує матрикс (строму) клітин. У гіалоплазмі є велика кількість ферментів, що беруть участь у процесах метаболізму, інформаційна і транспортна РНК. Через гіалоплазму здійснюється внутрішньоклітинний транспорт речовин. У ній накопичуються запасні поживні речовини. Органели — компоненти цитоплазми, які мають певну будову і спеціалізовану функцію. Їх поділяють на мікроскопічні та ультрамікроскопічні, загального призначення й спеціальні, мембранні та немембранні. Мікроскопічні органели видимі під світловим мікроскопом, а ультрамікроскопічні можна побачити за допомогою електронного мікроскопа. Органели загального призначення є в усіх клітинах, а спеціальні — в окремих різновидах клітин. Мембранні органели оточені клітинною мембраною, якої немає у немембранних органел. Мембранні органели являють собою частини цитоплазми, відмежовані від гіалоплазми клітинними мембранами. Вони мають специфічний для них вміст, який за своїми властивостями відрізняється від інших частин клітини. До мембранних органел належать мітохондрії, ендоплазматична сітка, комплекс Гольджі, лізосоми й пероксисоми. Мітохондрії — мікроскопічні органели загального призначення (рис. 5). В них утворюється енергія, яка накопичується у молекулах АТФ і необхідна для життєдіяльності клітин. Мітохондрії беруть також участь у регуляції обміну води, депонуванні йонів Са, Mg і P, продукуванні попередників стероїдних гормонів. Кількість мітохондрій у різних клітинах дуже варіабельна, від кількох до тисячі, що залежить від інтенсивності процесів метаболізму.
Найбільшу кількість мітохондрій виявлено в клітинах печінки, нервових клітинах, міоцитах та м’язових волокнах. Мітохондрії функціонують близько 20 діб, після чого утилізуються лізосомами. Нові мітохондрії утворюються шляхом поділу або брунькуванням. Під світловим мікроскопом мітохондрії мають вигляд нитко-, паличко- і зерноподібних структур завтовшки 0,2 – 2 мкм і завдовжки 1 – 10 мкм (див. рис. 5, а). Електронномікроскопічними дослідженнями встановлено, що мітохондрії обмежені двома мембранами завтовшки близько 7 нм (див. рис. 5, б). Між мембранами є проміжок завширшки 10 – 20 нм. Зовнішня гладенька мембрана відділяє їх від гіалоплазми, а внутрішня обмежує вміст мітохондрій — мактрикс. Вона утворює численні складки, спрямовані всередину мітохондрій, — кристи. На внутрішній поверхні внутрішньої мембрани розміщені мітохондріальні субодиниці грибоподібної форми, в яких є ферменти, що забезпечують процес фосфорилювання. В матриксі мітохондрій містяться ферменти циклу Кребса та окиснення жирних кислот. У матрикс із гіалоплазми надходять продукти обміну білків, жирів, вуглеводів, кисень і АДФ. Продукти обміну білків, жирів і вуглеводів у результаті окиснення розщеплюються до води та вуглекислого газу. Цей процес супроводжується виділенням енергії, що забезпечує фосфорилювання (приєднання до АДФ залишку фосфорної кислоти) і накопичується в молекулах АТФ, які залишають мітохондрії. У місцях використання енергії від молекул АТФ відщеплюється залишок фосфорної кислоти. При цьому виділяється вільна енергія, яка забезпечує життєдіяльність клітин. У матриксі мітохондрій є також автономний апарат для синтезу мітохондріальних білків. Він включає власне мітохондріальні ДНК, РНК, рибосоми та ферменти, необхідні для їх синтезу і синтезу білка. Однак цей апарат через недостатню інформацію не може забезпечити синтез усіх структур мітохондрій, і синтез більшості ферментів забезпечує геном ядра клітини.
Ендоплазматична сітка — ультрамікроскопічна органела загального призначення. Вона складається із системи видовжених цистерн, трубочок, канальців і мішечків, які сполучаються між собою. Ендоплазматична сітка є синтезуючою і транспортною системою клітини. На її мембрані синтезуються різні органічні речовини, компоненти клітинних мембран, які накопичуються в порожнинах елементів сітки і транспортуються в різні ділянки клітин. В останніх частини ендоплазматичної сітки з синтезованими речовинами відокремлюються і у вигляді пухирців надходять у гіалоплазму. Ендоплазматичну сітку поділяють на гранулярну і агранулярну. До зовнішньої поверхні мембран елементів гранулярної ендоплазматичної сітки прикріплюються у вигляді гранул рибосоми (рис..6). У зв’язку з цим функція цієї сітки полягає у синтезі білків як для потреб власне клітини, так і для виведення за її межі. Гранулярна ендоплазматична сітка дуже добре розвинена в клітинах, що синтезують білки. Зовнішня поверхня мембран елементів агранулярної ендоплазматичної сітки гладенька (див. рис. 6). У цій сітці відбувається синтез ліпідів і вуглеводів, детоксикація шкідливих продуктів обміну речовин, депонуванням йонів Са2+ (міоцити, м’язові волокна). Добре розвинена ця сітка в клітинах органів, які синтезують стероїдні гормони (надниркові залози, сім’яники).
|
|||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 185; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.138.141.202 (0.01 с.) |
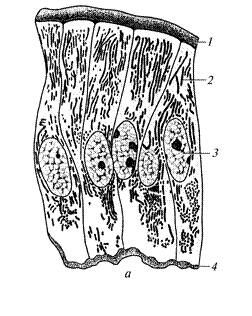

 Комплекс Гольджі — мікроскопічна органела загального призначення. За допомогою електронної мікроскопії встановлено, що до його складу входить чотири компоненти: транспортні пухирці, вигнуті цистерни, які розміщені одна над одною, конденсуючі вакуолі й секреторні гранули (див. рис. 6). У комплексі Гольджі відбувається накопичення, конденсація і дозрівання продуктів, синтезованих в ендоплазматичній сітці, синтез поліцукридів, здійснюється зв’язок поліцукридів з білками. Тут також синтезується надмембранний комплекс оболонки клітини, утворюються лізосоми й пероксисоми. В окремих різновидах клітин може бути від однієї до кількох десятків цих органел. Найбільша кількість комплексів Гольджі є в секреторних клітинах.
Комплекс Гольджі — мікроскопічна органела загального призначення. За допомогою електронної мікроскопії встановлено, що до його складу входить чотири компоненти: транспортні пухирці, вигнуті цистерни, які розміщені одна над одною, конденсуючі вакуолі й секреторні гранули (див. рис. 6). У комплексі Гольджі відбувається накопичення, конденсація і дозрівання продуктів, синтезованих в ендоплазматичній сітці, синтез поліцукридів, здійснюється зв’язок поліцукридів з білками. Тут також синтезується надмембранний комплекс оболонки клітини, утворюються лізосоми й пероксисоми. В окремих різновидах клітин може бути від однієї до кількох десятків цих органел. Найбільша кількість комплексів Гольджі є в секреторних клітинах.


