
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
ТемА: «ДиНаміка чисельності та її характеристика»Содержание книги
Поиск на нашем сайте
План: 1. Народжуваність, смертність, екологічні віки та виживання. 2. Щільність, чисельність та методи їх вимірювання. 3. Коливання чисельності - залежна та незалежна від щільності регуляція. 4. Стратегії популяцій.
Мета: Найбільш повно охарактеризувати народжуваність, смертність та виживання особин, які загалом визначають чисельність популяції.
1. Народжуваність – здатність популяції до збільшення. Застосування терміну до людини. Він характеризує частоту з`явлення нових особин в популяції, незалежно від способів розмноження (способи: ділення, брунькування, вилуплення, народження). Максимальна народжуваність (абсолютна або фізіологічна) – поява теоретичних максимальної кількості нових особин в ідеальних умовах (за відсутності лімітуючих факторів). Для даної популяції ця величина постійна. Екологічно реалізуєма народжуваність – означає збільшення популяції за фактичних умов середовища. Ця величина змінюється в залежності від розміру та складу популяції і фізичних умов середовища. Народжуваність виражають, як швидкість, визначеною шляхом ділення числа поновлених особин на час ( Народжуваність можна виміряти та виразити у різний спосіб:
N – показує або усю популяцію або її частину, здатну до розмноження. Наприклад: для вищих – в розрахунок на 1 самку. в (специфічна народжуваність) – для різних вікових груп або вікова. Наприклад: у ♀♀ дикого кролика розмір приплоду у віці 1-2 р., в середньому, перевищує 4 особини на 1 самку за рік, а у ♀♀ молодше року – 15 особин за рік. Народжуваність може бути нульовою але ніколи негативною. Але що стосується швидкості росту популяції, то Пояснити: різницю між приростом та народжуваністю. § Максимальна народжуваність дає приклад для співвідношення з розповсюдженою. Так, народжуваність в популяції миші 6 малюків на рік, буде мати більший задум, якщо відома межа до якої вона могла б збільшити в ідеальних умовах. § Є постійною величиною вона може бути використана для прогнозування швидкості збільшення популяції. Для практичної мети максимальна народжуваність встановлена експериментально. Наприклад: найвища середня продукція насінин люцерни за найбільш сприятливих умов вологості, температури та забезпеченості добривами – максимальна народжуваність. § Незвичне збільшення народжуваності або смертності викликає її тимчасові зміни, після чого виникає повернення до стабільного стану. Народжуваність відноситься до популяції, а не до окремих особин. За важелі народжуваності приймають середню, та аж ніяк не індивідуальну найбільшу або найменшу репродуктивну здатність. Окрім того, в деяких популяціях найвища швидкість розмноження за низької щільності, в інших за високої або середньої (приклад Оллі). Максимальна народжуваність проявиться у тому випадку коли не тільки відсутній лімітуючий вплив фізичних факторів, але й популяція має оптимальні розміри. По відношенню до розмноження розрізняють 3 екологічні віки: дорепродуктивний, репродуктивний, післярепродуктивний. Тривалість цих вікових категорій у тварин дуже сильно змінюється. У людини ці три вікові категорії приблизно однакові, на кожну припадає приблизно третина життя. Древня людина мала короткий післярепродуктивний період. У багатьох рослин і тварин дорепродуктивний період довгий, репродуктивний короткий, а післярепродуктивний відсутній. Наприклад: багатолітня сарана. Одноденки (від одного до декількох місяців). У сарани дуже довгий цикл (~17 років), але дорослі живуть менше одного сезону. Далекосхідні лососі, американська цикада Cicada sepemdecim – личинкова стадія має тривалість 17 років. Смертність характеризує загибель особин в популяції. Як прогнозують, її треба виразити числом особин, загиблих за даний період, або у вигляді специфічної смертності – по відношенню до числа особин які складають популяцію або ж її частину. Коефіцієнт смертності – d = dN ∕ dt – різниця, r = b - d – коефіцієнт приросту … популяції, або репродуктивний потенціал Наприклад: 100 особин – 10 померло: Екологічна смертність – загибель особин в даних умовах середовища, не постійна, залежить від стану популяції та екологічних факторів. Існує деяка теоретична мінімальна смертність – постійна величина характерна смертність в ідеальних умовах. Максимальна тривалість життя тільки тоді рівна фізіологічній тривалості і в середньому набагато перевищує екологічну тривалість життя. Зазвичай великий інтерес викликає не смертність, а – виживання. Якщо число загиблих особин = М, то тих, що вижили, = 1-М. Специфічну смертність показують як відсоток числа особин, загиблих за даний час від початкової чисельності популяції, або середньої чисельності популяції. Оскільки нас цікавить виживання, а не загиблі особини взагалі, доцільно показати смертність у вигляді величини зворотній виживанню. Як і у випадку народжуваності – необхідно знати: мінімальну величину смертності (теоретична константа), фактична або екологічна смертність. 2-у співставляємо з першою. В ідеальних популяціях вікова (старість) загибель? Тому мінімальна смертність визначається фізіологічною тривалістю життя. Оскільки у природі в більшості популяцій середня тривалість життя менша фізіологічну, то і смертність набагато перевищує мінімальну. Смертність як і народжуваність дуже сильно варіюється з віком. Чітке уявлення про смертність в популяції дають графіки їх вікової структури (вікові піраміди), таблиці про тривалість життя особин та криві виживання. Їх вивчення дозволяє судити в якому віці той або інший вид має вразливість. Якщо дія причин здатних змінити народжуваність або смертність припадає на найбільш слабку стадію, то її вплив на наступний розвиток популяції буде максимальним. Цей принцип використовується при влаштуванні мисливських господарств, і при боротьбі з шкідниками. В цілому фактори середовища більше впливають на смертність, аніж на народжуваність. Беручи до уваги дослідження популяції, варто звернути увагу на її щільність тому, що від величини щільності залежить вплив популяції на екосистему. Наприклад – один лось × 1000/ га лісу добре, але 20 чи 50 – це неймовірно! 2. Щільність популяції – це її величина по відношенню до одиниці простору. Вона вимірюється числом особин або біомасою, що припадає на одиницю площі або об’єму. Наприклад: 200 дерев/100 га, 19,8 зайців/100 га; 5 млн. діатомей/1 м 3 води; 200 кг риби/1 га водного дзеркала. Крім того одиницями біомаси можуть слугувати різні показники: від сухої ваги до вмісту ДНК або РНК. Інколи важливо проводити різницю між середньою щільністю (числом особин або біомасою за одиницю усього простору) і специфічною або екологічною щільність – число або біомаса за одиницю заселеного простору (тобто доступністю площі або об’єму, котрі можуть бути зайняті популяцією). Досить часто важливо знати збільшується або зменшується популяція, аніж її величину на даний момент. В таких випадках корисно використовувати показники відносної чисельності, які можуть відноситися до визначених проміжків часу. Наприклад: абсолютна кількість птахів, облікованих вдень або значення певних видів у відсотках (обліки на практиці з біології, екології). Наприклад: відсоток пробних ділянок, зайнятих астрагалом українським на схилах р. Молочної. Як і інші популяційні ознаки, щільність дуже мінлива. Однак ця мінливість не безмежна, існують верхні і нижні межі для популяції, які обмежують її розміри в природі протягом тривалого періоду. Приклад: На великій лісовій площі можуть жити, в середньому, 100 птахів/1 га; 20000 грунтових членистоногих /1 м2, але не буває 20000 птахів/1 м2, 10 членистоногих /1 га. Верхня межа щільності визначається потоком енергії в екосистемі (продуктивність), трофічним рівнем організму, а також величиною і інтенсивністю метаболізму. Нижня межа зазвичай не дуже помітна, але в стабільних екосистемах, де діють гомеостатичні механізми, підтримуючи щільність звичайних або панівних організмів з обмеженнями. Щільність популяції обмежується ресурсами двох типів: 1 – ресурси, що не поновлюються (простір або місце гніздування) 2 – ресурси, що поновлюються (їжа, вода, світло), якими популяція постачається безперервно. Потреби популяції можуть бути такими великими, що кількість ресурсів, що поновлюється, може впасти до нижньої межі. За таких умов, коли організмам стане важко їх добувати або асимілювати, подальший ріст популяції виявиться неможливим. Величина ресурсів, що поновлюються, зазвичай має незначні коливання завдяки збалансованості між їхньою експлуатацією та швидкістю відтворення. Щільність популяцій також дуже залежить від продуктивності екосистем. Наприклад:
Привести приклади: чим нижчий трофічний рівень тим більша щільність.
Якщо розміри і інтенсивність метаболізму відрізняються слабко, число особин слугує цілком задовільною мірою щільності, тому може досить часто трапитися така ситуація:
Загальне положення: Дані по чисельності призводять до перебільшення значення дрібних організмів, дані з біомаси – крупних. В той же час лише потік енергії слугує найбільш вірним критерієм для порівняння зазначених компонентів між собою. Одне з найбільших ускладнень при оцінюванні або вимірюванні щільності те, що досить часто особини в просторі розподілені нерівномірно і утворюють скупчення. Значимість вимірювання щільності величезна та має практичне значення: боротьба з шкідниками (комахи, гризуни), організація полювання, види, врожай, лікарські рослини тощо. Необхідно підкреслити, що показники відносної чисельності не варто змішувати з правильною щільністю, яка завжди відтворюється по відношенню до визначеного простору. Вище мова йшла про екологічну та середню щільність. Відмінність між ними, наприклад: 1) У національному лісовому парку Еверглейдс (США, Флорида) щільність дрібних риб зимою під час сухого періоду знижується у зв’язку з падінням рівня води, але екологічна щільність збільшується, тому що зі скороченням дзеркала води збільшується число риб, що приходиться на одиницю поверхні. Деякі птахи (лісовий лелека) припасовує відкладання яєць таким чином, що час вилуплення пташенят співпадає з піком екологічної щільності риб; це полегшує добування риби та годування малечі. Є багато методів вимірювання щільності популяції, які можна звести до 5-ти: 1) Загальний підрахунок, який застосовують для крупних та добре помітних організмів або для результатів їхньої діяльності (сайгаки, вівцебик, північний олень, ондатра, бобер). Щільності у птахів. Прямий підрахунок застосовують і для комах. Фр. еколог Дрьо вважає, що середня щільність комах в Альпах – 2 особини на 1 м 2. 2) Метод пробних ділянок – підрахунок або зважування організмів на відповідність числа ділянок або трансектів відповідних розмірів для оцінки щільності на дослідницькій площі. 3) Метод мічення з наступним відловом (для рухомих тварин), а) мічення різними способами (кільця; вушні, шийні мітки; тавро рідким азотом), б) відлов. Приклад: Було тавровано 100 тварин; серед пійманих 100 особин - лише 10 мали тавро.
4) Метод вилучення – при якому число організмів які вилучаються з деякої площі відмічають на осі ординат, а число вилучених раніше з цієї ж території – на осі абсцис. Якщо вірогідність відлову відносно постійна, то крапки розташовуються по прямій 5) Безділянковий метод – метод найближчого сусіди. Щільність за відстанню на одиницю площі. Приклади. Підрахунок кількості дерев у лісі. Підрахунок кількості місць мешкання певних тварин та самих тварин. 3. Коливання чисельності. Народження, смертність, вікова структура самі собою важливі, але кожен з них окремо дуже мало, що пояснюють показником, котрий узагальнює їх – є ріст популяції. Для позначення максимальної репродуктивної здатності існує термін "біотичний потенціал" (Чентен, 1928 р.). Ця природжена властивість організму до розмноження та виживання тобто до збільшення чисельності; це свого роду алгебрична сума числа молодих особин, виробляючи за кожен цикл розмноження, число репродукцій за даний проміжок часу, співвідношення статі та загальною здатністю до виживання за даних фізичних умов. Збільшення чисельності популяції виникає не шляхом простого додавання (арифметичний зріст), а множення (геометричний зріст). Геометрична швидкість росту популяцій в ідеальних умовах виявляють важливу обставину: популяції володіють величезною внутрішньою властивою здатністю до росту. Якщо б цей зріст не утримувався обмеженістю необхідних ресурсів або хижаками і паразитами, то чисельність більшості видів досягала б астрономічних цифр. Ч. Дарвін "Походження видів…", "не існує жодного винятку з правил за яким будь-яка органічна речовина природно розмножується у настільки швидкій прогресії, що не піддається його винищенню, нащадки однієї б пари заполонили б усю земну кулю". Для того, щоб повірити в це необхідно простежити за збільшенням чисельності популяції за інтродукції виду в новий район. Наприклад: § Свійська вівця. Тасманія. 1820 р. – 200 тис. особин; 1850 р. ~ 2 млн. особин. Десятикратне збільшення за 30 років еквівалентно щорічному збільшенню на 8%. § Звичайний фазан. Острів Протекшен (штат Вашингтон) 1937 р. – 20 +6. Через 5 років ~ 1325 особин. Це збільшення чисельності у 166 разів відповідає щорічному збільшенню на 180%. § Каліфорнійський морський лев був майже винищений в XIX ст.. мисливцями: у 1890 р. ~ 20 особин. Охорона популяції. 1970р. – 30000. За 100 років без обмежень їх чисельність могла б сягнути 27 млн. особин. Високі швидкості росту популяції зручно виражати в часі необхідному для подвоєння чисельності. Так для морського слона воно дорівнює ~7,6 років, для фазана ~ 8 місяців, для нориці ~ 80 днів, для мучного хрущака ~ 10 днів і для дафнії ~ 3 дні. Характер збільшення чисельності популяції може бути різним. У зв’язку з цим виокремлюють різні типи росту. По формі кривих, побудованих на арифметичній шкалі можна показати 2 основні: ріст, описаний І-подібною і S-подібною кривою. Ці два протилежних типи росту у відповідності з особливостями різних організмів та умов середовища можуть поєднуватися і модифікуватися. При І-подібній щільність швидко збільшується, але потім, коли починає діяти протистояння середовища або інший лімітований фактор (більш або менш різко), ріст популяції припиняється.
А-флуктуації зазвичай не регулярні і обумовлені впливом одного або кількох лімітуючих факторів (опади, t0). Інші б-флуктуації настільки регулярні, що їх можна назвати циклічними. Різкі осциляції характерні для популяцій з – І-подібним типом росту, а загасаючи – з S-подібним. Роз’яснення механізмів, які лежать в основі змін чисельності популяцій, – дуже важлива екологічна проблема, котра ще повністю не розв’язана. Всім добре відомі сезонні зміни (флуктуації) величини популяції. Наприклад: комарі, амброзія, зайці, комахи тощо. Хоча у природі важко знайти популяції організмів, у яких би не виникали зміни за сезонами, найбільш виражені флуктуації відмічаються у організмів, які обмежені періодом розмноження та коротким життєвим циклом або зі значними сезонними розповсюдженнями у просторі, наприклад у мігруючих тварин. У таких організмів найчастіше: J-подібні криві чисельності. Сезонна динаміка чисельності шпаків, жаб, зайців, кабанів тощо за рахунок молодняку. Приріст. У тропіках флуктуації найчастіше пов’язані з періодом дощів, але вони можуть бути узагальнені різного роду періодичністю у співтоваристві. Наприклад, у комах та птахів, щільність може коливатися в залежності від періодичності квітування та плодоношення рослин, що мають місце у лісах навіть за відсутності періодичних дощів. Різкий спалах чисельності популяцій якій виявився у новому місці популяцій – лось – на острові Ньюфаундленд, ондатра – у Європітощо. Розрізняють також неперіодичні та рідко спостережні коливання чисельності. Наприклад: § Американський оселедець (Alosa sapidissima). В 1900 р. щорічний вилов ~ 2000 т. Інтенсивний вилов, спорудження гребель, що перешкоджають нересту, знизили видобуток на 98%. Здавалося, що вид зникне назовсім, але раптово досяг високої чисельності. В 1944 р. – вилов оселедцю склав ~ 2500т. Зараз чисельність знову пішла на зниження. § Зміни у виловах скумбрії у берегів Північної Америки з 1884 по 1886 рр. вилов впав від 60 000 т. до 10 000 т. на рік. § З 1966 р. – ріст чисельності морської зірки Acantpaster pianci. Був вид мало чисельний – < 1особина/1м2, зараз = 1/1м2. Харчується поліпами коралових рифів. Очистила 40 км- смугу рифів навколо острова Гуам менш ніж за 3 роки. Скелет коралів легко руйнується від ерозії. Покидають риби, руйнуються атоли. Жодна з гіпотез (ДДТ, зникнення його основи корму) не підтвердилася. Черевоногі молюски Charonia fritonis – добувають для перламутру; радіоактивні опади…
|
|||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-05; просмотров: 396; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.26.149 (0.012 с.) |

 N і
N і  = В (народжуваність).
= В (народжуваність).  = в, або народжуваність за одиницю популяції.
= в, або народжуваність за одиницю популяції.
 × 100.
× 100.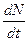 =rN визначеною межею для N, r - швидкість росту.
=rN визначеною межею для N, r - швидкість росту.
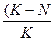 – Верхня межа збільшення чисельності має назву межа росту.
Для природних популяцій важливо розрізняти:
1) сезонні зміни їхньої величини, котрі регулюються головним чином онтогенетичними адаптаціями, залежними від сезонних змін фактору середовища, 2) річні флуктуації –
а) обумовлені різницею фізичних факторів середовища протягом року;
б) осциляції, обумовлені внутрішніми змінами у популяції, які відбуваються внаслідок внутрішньовидової конкуренції, від накопичення старих особин тощо.
– Верхня межа збільшення чисельності має назву межа росту.
Для природних популяцій важливо розрізняти:
1) сезонні зміни їхньої величини, котрі регулюються головним чином онтогенетичними адаптаціями, залежними від сезонних змін фактору середовища, 2) річні флуктуації –
а) обумовлені різницею фізичних факторів середовища протягом року;
б) осциляції, обумовлені внутрішніми змінами у популяції, які відбуваються внаслідок внутрішньовидової конкуренції, від накопичення старих особин тощо.
 Рис. Динаміка чисельності козулі (1) і її приросту (2)
у степових популяціях України.
Рис. Динаміка чисельності козулі (1) і її приросту (2)
у степових популяціях України.



