Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Корреляции разных типов родственников
Типы родственников VA VD VEC Биологические родители и дети 1/2 VC(BPO) Приемные родители и дети VС(АРО) Сиблинги с одним общим родителем 1/4 C C(HS) Сиблинги 1/2 1/4 VС(FS) Двуяйцевые близнецы 1/2 1/4 VC(DZ) Однояйцевые близнецы 1,0 1,0 VC(MZ) Примечание. Здесь и далее: ВРО — родители Х дети (biological parent-offspring); APO — приемные родите- ли Х дети (adopted parent-offspring); HS — полусиблинги (half-sibling); FS — пол- ные сиблинги (full-sibling); DZ— ДЗ близнецы (dizygotic twins); MZ — МЗ близ- нецы (monozygotic twins). С целью максимизации информации, полученной при анализе раз- ных типов родственников, ученые совмещают несколько методов в рам- ках одного исследования. Выбор методов для исследования того или иного признака является специальной задачей. Главное правило здесь заключается в том, что количество независимых исходных статистик (т.е. количество корреляций между родственниками) должно превышать количество неизвестных в системе уравнений. Если это правило не выдерживается, система уравнений однозначного решения не имеет. Например, представим себе, что мы исследуем по некоторому признаку биологические семьи, каждая из которых растит по крайней мере двух детей. Соответственно, мы можем определить корреляции по исследуемому признаку как между родителями и детьми, так и между сиблингами в данных семьях. Любая из этих пар будет иметь в среднем 50% общих генов, что позволяет, используя информацию из табл. 8.5, записать следующую систему уравнений: r = [1 / V + V ]/ V, BPO A C (BPO) P
r = [1 / V + / V + V ]/ V. FS A D C (FS) P
Очевидно, что полученное после некоторых алгебраических пре- образований уравнение r − V = r −[1/ V + V ]/ V, одно- BPO C (BPO) FS D C (FS) P значного решения не имеет, поскольку в нем присутствуют только 2 известных, но 3 неизвестных члена. Напротив, совмещение родительско-детских корреляций, полу- ченных в приемных и биологических семьях, позволяет записать сис- тему уравнений, решающих эту проблему: r = [1/ V + V ]/ V, BPO A C (BPO) P r = V / V. APO C (APO) P После преобразования получим r − / V = r. BPO A APO Данное уравнение имеет однозначное решение, поскольку V / V = ( 2 r − r). A P BPO APO Чем больше различных пар родственников включено в анализ, тем больше компонентов дисперсии может быть определено однозначно и
тем более сложные и разветвленные модели могут оцениваться. В качестве иллюстрации рассмотрим два метода, используемых для разделения генетической и средовой составляющих фенотипической дисперсии в популяции (подробнее о методах психогенетики — в гл. VII). Метод близнецов. Этот метод, без сомнения, был и до сих пор является одним из ведущих методов психогенетики. Классический ва- риант метода близнецов основывается на том, что монозиготные (МЗ) и дизиготные (ДЗ) близнецы характеризуются различной степенью генетического сходства, в то время как их среда может считаться при- близительно одинаковой. На языке составляющих фенотипической дисперсии (см. табл. 8.2 и 8.3) это можно выразить так: r = (V + V + V) / V; MZ A D C P r = (1 / V + / V + V) / V; DZ A D C P I = (V + V + V + V) / V. A D C N P Соотнеся первое и третье1 − r = V / V. уравнения, получим: NZ N P МЗ близнецы представляют собой идентичные генетические ко- пии друг друга, поэтому теоретически корреляция МЗ близнецов по признаку, вариативность которого в популяции находится полностью под генетическим контролем, должна равняться 1,0. Разницу между 1,0 и реальной корреляцией МЗ близнецов можно объяснить влияни- ями индивидуальной среды или ошибки измерения (компонент VN содержит в нерасчлененном виде обе эти составляющие). Отметим, что приведенные закономерности соотношения МЗ и ДЗ близнецов справедливы только при следующих условиях (частично речь о них шла в гл. VII):
1. Центральным допущением при использовании метода близне- цов в любом его варианте является допущение о равенстве среды МЗ и ДЗ близнецов. Важно отметить, что оно подразумевает не одинако- вость близнецовых сред, а тот факт, что распределение (частота встре- чаемости и разброс) средовых компонентов монозиготных близнецов не превышает разнообразия сред дизиготных. Правомерность этого допущения до сих пор исследуется и обсуждается психогенетиками; если оно не справедливо, то получаемые этим методом оценки коэф- фициента наследуемости искажены. Как уже говорилось, это допуще-
ние касается не всей близнецовой среды, а только тех ее аспектов, которые связаны с изучаемым признаком (если они известны). 2. V GxЕ = 0, т.е. принимается допущение об отсутствии ГС-взаи- модействия. Заметим, что в некоторых случаях такое допущение впол- не правомерно, в большинстве же случаев оно требует тщательной эмпирической проверки. 3. Cov(G)(E) = 0, т.е. принимается допущение об отсутствии геyотип- средовой ковариации. Прямо проверить это допущение в рамках клас- сического близнецового метода невозможно. Поэтому, как и в случае двух предыдущих допущений, отсутствие ГС-ковариации и корреля- ции при использовании классического метода близнецов принимает- ся на веру. 4. Ассортативность по исследуемому признаку не отличается от нуля (т.е. μ = 0). Как уже говорилось, это допущение для большинства исследуемых в психогенетике признаков неверно: неслучайность под- бора супружеских пар у человека — скорее правило, чем исключение. Поэтому допущение об отсутствии ассортативности надо обязательно проверять (в том случае, если в литературе отсутствуют необходимые сведения) по данным о супружеских парах. В общем случае корреля- ция между супругами включает в себя компонент, обусловленный ассортативностью брака, и компонент, обусловленный влиянием се- мейных систематических средовых факторов. Самым простым и на- дежным способом проверки этого допущения является обследование родителей близнецов. Не имея данных о родителях (т.е. корреляций между родителями по исследуемому признаку), исследователь не мо- жет «развести» эффекты ассортативности и эффекты семейной среды. Наличие же значимой ассортативности повышает возможность полу- чения ДЗ одинаковых генов от обоих родителей (у МЗ и без этого фактора их 100%), повышая r ДЗ и тем самым снижая разность rМЗ - rДЗ и, следовательно, величину коэффициента наследуемости (о нем речь пойдет ниже). 5. В генетическом механизме изучаемого признака отсутствуют эпи- статические взаимодействия (V t ). Это условие принимается как долж- ное практически во всех психологических исследованиях (многие ис- следователи принимают данное допущение a priori, даже не обсуждая его правомерность). Однако в ситуациях, когда это допущение не-
справедливо, оценки составляющих фенотипической дисперсии мо- гут быть сильно искажены, поскольку эпистатическое взаимодействие генов может значительно уменьшить генетическое сходство ДЗ близ- нецов, тем самым увеличивая разницу между r МЗ и r ДЗ и приводя к завышенным оценкам коэффициента наследуемости. Однако даже в том (весьма неправдоподобном!) случае, когда ис- следуется психологический признак, для которого соблюдаются все вышеперечисленные условия, оценить все четыре компонента феноти- пической дисперсии (V A,VD,VC,VN) в рамках метода близнецов невоз- можно, так как четыре независимых величины не могут быть определе- ны из трех линейных уравнений. Ученые, тем не менее, сделав несколь- ко упрощающих допущений, разработали несколько способов оценки коэффициента наследуемости на основе метода близнецов. Отметим, что ни один из этих методов не является «правильным» или «непра- вильным» — каждый из них обладает определенными достоинствами
и недостатками. Рассмотрим кратко хотя бы три наиболее часто встре- чающихся в литературе метода оценки коэффициента наследуемости. КОЭФФИЦИЕНТ ХОЛЬЦИНГЕРА К. Хольцингер предложил следующую формулу для оценки насле- дуемости: r − r V + / V MZ DZ A D H = = . − r V + / V + 2 V DZ A D N Данная формула, как и следующая, адекватна только в случае, если среда МЗ равна таковой у ДЗ, при наличии же VD эта оценка будет смещенной. Поскольку в этой формуле VС и VN заменены на удвоенную V N, то нетрудно заметить, что при VC < VN этот коэффи- циент будет завышен, а при VC > VN, наоборот, занижен. КОЭФФИЦИЕНТ ИГНАТЬЕВА* В качестве первой оценки величины генетической составляющей фенотипической дисперсии часто используется коэффициент Игна- тьева, вычисляемый следующим образом:
* Данный способ оценки генетического компонента дисперсии и зарубежной психогенетике связан с именем Д. Фальконера, работа которого вышла в I960 г. Однако этот коэффициент был предложен еще в 1934 г. М.В. Игнатьевым. Кратко об этом см. во Введении, а также в работах В.М. Гиндилиса [97] и Б.И. Кочубея [132, гл. I]. В формуле Игнатьева используются иные символы, но, поскольку в современной науке утвердились приводимые далее обозначения, будем пользо- ваться ими и мы. В приводимой ниже формуле Еобщ — то же, что ЕС, a Eинд — то же, что EN в предыдущем тексте (см. табл. 8.3).
V + V / h = ( 2 r − r) A D = . MZ DZ VP При наличии доминантного компонента дисперсии VD оценка наследуемости будет завышена. Очевидно, что влияние любых факторов, изменяющих разницу между корреляциями двух типов близнецов (например, завышение корреляции между МЗ близнецами, возникающее в результате дей- ствия специфической для этого типа близнецов среды), будет влиять на эту оценку наследуемости. Хотя в последние годы появились и все чаще употребляются более современные и сложные методы статисти- ческого анализа, этот коэффициент, в силу своей аргументированно- сти и простоты получения, остается в арсенале психогенетики. Более того, Р. Пломин предложил с помощью этой формулы оценивать — тоже в первом приближении, конечно, — и долю средовых компо- нентов: E = r − h, E = 1 − h − E, общ MЗ инд общ где Eoбщ — общесемейная среда (VС), Еинд — индивидуальная среда (VN). Правда, в оценку индивидуальной среды неизбежно включается часть дисперсии, вызванная ошибкой измерения. Возможность кор-
рекции этого дефекта обсуждена выше. МЕТОД ДЕ ФРИЗА И ФУЛКЕРА (ДФ-МЕТОД) Дж. де Фриз и Д. Фулкер разработали две регрессионные модели: 1) классическую регрессионную модель, в которой частная регрессия значения со-близнеца на значение близнеца—условного пробанда и коэффициент родства представляет собой тест генетической этиоло- гии исследуемого признака, и 2) расширенную регрессионную мо- дель, предоставляющую прямое свидетельство того, насколько инди- видуальные различия внутри исследуемой группы объясняются гене- тическими и средовыми влияниями. Эти два регрессионных уравнения записываются следующим образом: C = B P + B R +; A
C = B P + B R + B PR +, A где С — значение со-близнеца по исследуемому признаку (данный метод подразумевает выделение в каждой паре одного близнеца — условного пробанда, тогда второй близнец называется со-близнецом); Р — значение близнеца-пробанда по тому же признаку; R — коэффи- циент родства (1 для МЗ и 0,5 для ДЗ близнецов); PR — произведение
значения пробанда по исследуемому признаку на коэффициент род- ства; А — константа регрессионного уравнения. Решение этих уравнений позволяет оценить следующие парамет- ры: В1, представляет собой показатель среднего сходства между МЗ и ДЗ близнецами; В2 — оценку удвоенной разницы между средними в группах МЗ и ДЗ близнецов (с учетом ковариации между значениями МЗ и ДЗ пробандов); В3 оценивает долю дисперсии, объясняемую сре- довыми влияниями, общими для членов близнецовой пары (VС /VР или С2); В4, отражает разницу h2g - h 2, где h2 — коэффициент наследу- емости в широком смысле и h2g — коэффициент наследуемости в оп- ределенной группе (например, коэффициенты наследуемости IQ в группах здоровых людей и людей, страдающих ФКУ, отличаются друг от друга; В4 показывает разницу коэффициентов наследуемости, по- лученных в генеральной популяции и специфической выборке); и, наконец, В5 оценивает коэффициент наследуемости (h2), т. е. показа- тель того, насколько индивидуальные различия в исследуемой выбор- ке объясняются наследуемыми влияниями. Интересной особенностью ДФ-метода является то, что он позво- ляет тестировать гипотезу о сходстве или различии этиологии нор- мально распределенных и экстремальных значений. Сравнение рег- рессионных коэффициентов В2 и В5 позволяет проверить гипотезу о том, сходны ли этиологии девиантных и «средних» значений, напри- мер, по тесту на математические способности. Если этиология неспо- собности к математике отличается от этиологии средних математи- ческих способностей, то В2 и В5 должны статистически надежно отли- чаться друг от друга. Если же дети, которые имеют трудности в овладении математикой, представляют собой не отдельную группу, а край нормального распределения, то В2 и В5 статистически отличать- ся друг от друга не должны, Разные формулы для вычисления коэффициентов наследуемости характеризуются разного рода допущениями и ограничениями. В не-
скольких исследованиях было продемонстрировано, что применение разных формул на одном и том же эмпирическом материале дает раз- ные результаты. Поэтому интерпретация данных, полученных одним методом близнецов, должна проводиться с учетом всех ограничений, свойственных этому методу. Ф. Фогель и А. Мотульски [159] отмечают, что даже при сильно упрощающих допущениях (например, отсутствия ассортативности, доминирования и т.д.) все равно остаются система- тические ошибки, которые невозможно полностью проконтролиро- вать. Они рекомендуют вычислять из одних и тех же эмпирических данных альтернативные оценки и сравнивать, насколько хорошо они совпадают. Метод приемных детей. При допущении, что среда семей-усыно- вителей не коррелирует со средой тех биологических семей, из кото- рых данные дети усыновляются, корреляции детей с их биологичес-
кими родителями представляют собой «чистые» генетические корре- ляции (т.е. прямую оценку h2 или VG /VP, а с родителями-усыновите- лями — «чистые» средовые корреляции (с2 или V С /VP). Однако в том случае, если среды биологических и приемных семей похожи, допу- щение о «чистоте» полученных оценок генетической и средовой со- ставляющих чаще всего неправомерно (по крайней мере в тех случа- ях, когда корреляция сред неизвестна). Методологически адекватным, хотя практически и не всегда возможным решением в подобной ситу- ации служит получение нескольких оценок генетического и средово- го компонентов при разных значениях корреляции сред. Таким образом, главной причиной беспокойства при использова- нии метода приемных детей является допущение об отсутствии кор- реляции между биологическими и приемными семьями. Кроме того, исследователи должны убедиться в том, что семьи-усыновители реп- резентативны общей популяции, т.е. не отличаются от среднепопуля- ционной семьи по уровню благосостояния, образования и т.п. Если семьи-усыновители нерепрезентативны, закономерности, полученные в результате их анализа, не могут считаться справедливыми для гене- ральной популяции. АНАЛИЗ ПУТЕЙ Приведенная выше логика разложения фенотипической диспер- сии на ее составляющие, реализованная в нескольких эмпирических методах, представляет собой один из способов определения коэффи- циента наследуемости того или иного признака. Но понятие наследу- емости можно также проанализировать при помощи «анализа путей». Анализ путей в последние десятилетия широко используется и в психогенетике, и в науках о поведении вообще. Он был предложен генетиком С, Райтом еще в 30-х годах и затем им же и другими иссле- дователями детально разработан. Четкое изложение его основ и пра- вил использования содержится в упоминавшемся труде М. Нила и Л. Кардона [342], которые характеризуют этот метод следующим образом. Диаграмма путей — эвристичный способ наглядного графическо- го представления причинных и корреляционных связей (путей) меж- ду переменными, позволяющий дать полное математическое описа- ние линейной модели, которую применяют исследователи. Тем са- мым диаграмма путей способствует ее пониманию, верификации или представлению результатов. В целом путевые модели — «экстремально обобщенный» способ анализа, один из многих мультивариативных методов (к ним же относятся методы множественной регрессии, фак- торный и дискриминантный анализы и т.д.). Существуют определенные правила построения диаграмм пу- тей (рис. 8.4). Прямоугольники (или квадраты) обозначают наблюда-
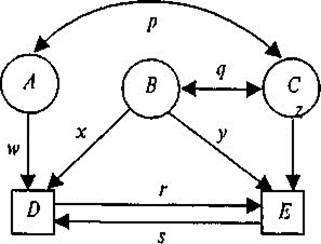
Рис. 8.4. Диаграмма путей, объединяющая три латентных (А, В, С) и две наблюдаемых (D и Е) переменных. р и q — корреляции; r, s, w, х, у, z — путевые коэффициенты.
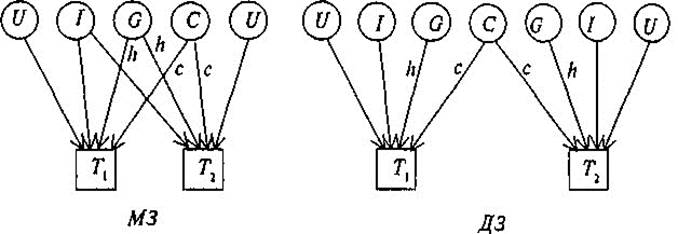
Рис. 8.5. Диаграмма путей для корреляций совместно живущих пар МЗ и ДЗ близнецов. Т1, Т2 — близнецы одной пары. G — генотип; С— общая среда; U — индивидуаль- ная (уникальная) среда; I— эпистаз. Пути h, с — влияния G, С на исследуемую черту. емые переменные; круги (или эллипсы) — латентные, неизмеряе- мые переменные (на рис. 8.4. D и Е; А, В, С соответственно). Связи между переменными обозначаются стрелками: постулиро- ванные исследователем причинно-следственные — направленной в одну сторону («путь» от причины к следствию); наблюдаемые ассо- циации — двусторонней. На рис. 8.4 первые — w, x, у, z, r, s (путевые коэффициенты); вторые — р и q (коэффициенты корреляции). Ина- че говоря, модель выделяет зависимые переменные (D и Е), подле- жащие объяснению или прогнозированию, и независимые (А, В, С), действие которых должно объяснить или предсказать зависимые пе- ременные и их связи. Есть и другие, более детальные, правила офор- мления и чтения путевых диаграмм, но мы их рассматривать не будем. На рис. 8.5 даны модели путей для корреляций совместно живу- щих пар МЗ и ДЗ близнецов по экстраверсии, из которых следует, что
корреляция МЗ близнецов T1 и Т2 может быть выражена через сумму путей, связывающих их, т.е. hh и сс; иначе говоря, r МЗ = h2 +с2. Для ДЗ это будут пути h х 1/2 х h и cc, т.е. rДЗ = 1/2 h2 + с2. Вычитая, получим rМЗ — rДЗ = h2 + с2 — 1/2 h2 — с2 = 1/2 h2; чтобы получить полную генетичес- кую дисперсию (а не половину ее), удваиваем разность корреляций h2 = 2(rMЗ — rДЗ) и получаем описанный выше коэффициент наследу- емости, справедливый для близнецовых исследований. Аналогичным образом могут быть построены путевые диаграммы для семейных и любых других данных. Единицы измерения, используемые в анализе путей, отличаются от тех, которыми мы оперировали тогда, когда рассматривали по- нятие наследуемости на примере разложения фенотипической дис- персии. Если при разложении дисперсии мы пользовались квадратич- ными единицами (например, h2, V G), то в данном случае наследуе- мость описывается на языке стандартных отклонений. Тогда путевые коэффициенты являются коэффициентами регрессии, полученными для переменных не в исходных единицах, а для стандартизованных переменных. Несмотря на широкое использование этого метода и его достоин- ства, которые заключаются прежде всего в наглядной демонстрации представлений о компонентах, влияющих на исследуемый признак, он имеет и своих критиков. Так, Ф. Фогель и А. Мотульски «не уверены в том, что этот метод биометрического анализа внесет существенный вклад в наше понимание генетических факторов» [159]. Одно из глав- ных сомнений вызывает тот факт, что в диаграмму путей и, следова- тельно, в дальнейший математический анализ закладываются уже имеющиеся у исследователя предположения о влияющих на признак факторах, их причинно-следственных отношениях и т.д., и результат анализа зависит, таким образом, от корректности заранее имеющих- ся исходных позиций.
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-01-26; просмотров: 258; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.161.116 (0.163 с.) |