Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Типы и режимы мышечных сокращенийСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
− В зависимости от условий, в которых происходит мышечное сокращение, различают два его основных типа — изотоническое и изометрическое. Сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается постоянным, называется изотоническим. Изометрическим является такое сокращение, при котором мышца укоротиться не может, если оба ее конца закреплены неподвижно. В этом случае по мере развития сократительного процесса напряжение возрастает, а длина мышечных волокон остается неизменной. В натуральных двигательных актах сокращения мышц смешанные: даже поднимая постоянный груз, мышца не только укорачивается, но и изменяет свое напряжение вследствие реальной нагрузки. Такое сокращение называется ауксотоническим. − В зависимости от частоты стимуляции выделяют одиночные и тетанические сокращения. Одиночное сокращение (напряжение) возникает при действии на мышцу одиночного электрического или нервного импульса. Волна возбуждения возникает в месте приложения электродов для прямого раздражения мышцы или в области нервно-мышечного соединения и отсюда распространяется вдоль всего мышечного волокна. В изотоническом режиме одиночное сокращение икроножной мышцы лягушки начинается через короткий скрытый (латентный) период — до 0,01 с, далее следуют фаза подъема (фаза укорочения) — 0,05 с и фаза спада (фаза расслабления) — 0,05—0,06 с. Обычно мышца укорачивается на 5—10 % от исходной длины. Как известно, продолжительность волны возбуждения (ПД) мышечных волокон варьирует, составляя величину порядка 1—10 мс (с учетом замедления фазы реполяризации в ее конце). Таким образом, длительность одиночного сокращения мышечного волокна, наступающего вслед за его возбуждением, во много раз превышает продолжительность ПД. Мышечное волокно реагирует на раздражение по правилу «все или ничего», т.е. отвечает на все надпороговые раздражения стандартным ПД и стандартным одиночным сокращением. Однако сокращение целой мышцы при ее прямом раздражении находится в большой зависимости от силы стимуляции. Это связано с различной возбудимостью мышечных волокон и разным расстоянием их от раздражающих электродов, что ведет к неодинаковому количеству активированных мышечных волокон. При пороговой силе стимула сокращение мышцы едва заметно, потому что в реакцию вовлекается лишь небольшое количество волокон. При увеличении силы раздражения число возбужденных волокон растет, пока все волокна не окажутся сокращенными, и тогда достигается максимальное сокращение мышцы. Дальнейшее усиление стимулов прироста амплитуды сокращения не вызывает.
В естественных условиях мышечные волокна работают в режиме одиночных сокращений только при относительно низкой частоте импульсации мотонейронов, когда интервалы между последовательными ПД мотонейронов превышают длительность одиночного сокращения иннервируемых ими мышечных волокон. Еще до прихода следующего импульса от мотонейронов мышечные волокна успевают полностью расслабиться. Новое сокращение возникает после полного расслабления мышечных волокон. Такой режим работы обусловливает незначительную утомляемость мышечных волокон. При этом ими развивается относительно небольшое напряжение.
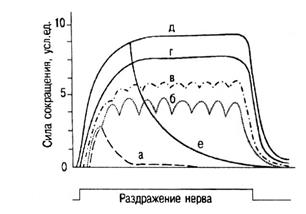
Тетаническое сокращение — это длительное слитное сокращение скелетных мышц. В его основе лежит явление суммации одиночных мышечных сокращений. При нанесении на мышечное волокно или целую мышцу двух быстро следующих друг за другом раздражении возникающее сокращение будет иметь большую амплитуду. Сократительные эффекты, вызванные первым и вторым раздражениями, как бы складываются, происходит суммация, или суперпозиция, сокращений, поскольку нити актина и миозина дополнительно скользят друг относительно друга. При этом в сокращение могут вовлекаться ранее не сокращавшиеся мышечные волокна, если первый стимул вызвал у них подпороговую деполяризацию, а второй увеличивает ее до критической величины. При получении суммации в одиночном волокне важно, чтобы второе раздражение наносилось после исчезновения ПД, т.е. после рефрактерного периода. Естественно, что суперпозиция сокращений наблюдается и при стимуляции моторного нерва, когда интервал между раздражениями короче всей длительности сократительного ответа, в результате чего и происходит слияние сокращений. При сравнительно низких частотах наступает зубчатый тетанус, при большой частоте — гладкий тетанус (рис. 13).
Рис. 13. Сокращения икроножной мышцы лягушки при увеличении частоты раздражения седалищного нерва. Суперпозиция волн сокращения и образование разных видов тетануса. а — одиночное сокращение (Г = 1 Гц); б, в — зубчатый тетанус (Г= 15—20 Гц); г, д — гладкий тетанус и оптимум (Г = 25—60 Гц); е — пессимум — расслабление мышцы во время раздражения (Г= 120 Гц).
Их амплитуда больше величины максимального одиночного сокращения. Напряжение, развиваемое мышечными волокнами при гладком тетанусе, обычно в 2—4 раза больше, чем при одиночном сокращении. Режим тетанического сокращения мышечных волокон в отличие от режима одиночных сокращений быстрее вызывает их утомление и поэтому не может поддерживаться длительное время. Из-за укорочения или полного отсутствия фазы расслабления мышечные волокна не успевают восстановить энергетические ресурсы, израсходованные в фазе укорочения. Сокращение мышечных волокон при тетаническом режиме с энергетической точки зрения происходит «в долг». До сих пор нет общепризнанной теории, объясняющей, почему напряжение, развиваемое при тетанусе, или суперпозиции сокращений, гораздо больше, чем при одиночном сокращении. Во время кратковременной активации мышцы вначале одиночного сокращения в поперечных мостиках между нитями актина и миозина возникает упругое напряжение. Однако недавно было показано, что такой активации недостаточно для прикрепления всех мостиков. Когда она более длительная, обеспечиваемая ритмической стимуляцией (например, при тетанусе), их прикрепляется больше. Количество поперечных мостиков, связывающих актиновые и миозиновые филаменты (а, следовательно, и развиваемая мышцей сила), согласно теории скользящих нитей, зависит от степени перекрывания толстых и тонких нитей, а значит, и от длины саркомера или мышцы. Высвобождение Са2+ при тетанусе. Если стимулы поступают с высокой частотой (не менее 20 Гц), уровень Са 2 + в интервалах между ними остается высоким, потому что кальциевый насос не успевает вернуть все ионы в продольную систему саркоплазматического ретикулума. В таких условиях отдельные сокращения почти полностью сливаются. Это состояние устойчивого сокращения, или тетанус, наблюдается в том случае, когда промежутки между стимулами (или потенциалами действия в клеточной мембране) меньше примерно 1/3 длительности каждого из одиночных сокращений. Следовательно, частота стимуляции, необходимая для их слияния, тем ниже, чем больше их длительность; по этой причине она зависит от температуры. Минимальный промежуток времени между последовательными эффективными стимулами во время тетануса не может быть меньше рефрактерного периода, который приблизительно соответствует длительности потенциала действия. Как выяснилось, амплитуда гладкого тетануса колеблется в широких пределах в зави-\ симости от частоты стимуляции нерва. При некоторой оптимальной (достаточно высокой) частоте стимуляции амплитуда гладкого тетануса становится наибольшей. Такой гладкий тетанус получил название оптимума. При дальнейшем повышении частоты стимуляции нерва развивается блок проведения возбуждения в нервно-мышечных синапсах, приводящий к расслаблению мышцы в ходе стимуляции нерва — пессимум Введенского. Частота стимуляции нерва, при которой наблюдается пессимум, получила название пессимальной (см. рис. 6.4).
В эксперименте легко обнаруживается, что уменьшенная в ходе пессимальной ритмической стимуляции нерва амплитуда мышечного сокращения моментально возрастает при возвращении частоты раздражения от пессимальной к оптимальной. В этом наблюдении — хорошее доказательство того, что пессимальное расслабление мышцы не является следствием утомления, истощения энергоемких соединений, а является следствием особых соотношений, складывающихся на уровне пост- и пресинаптических структур нервно-мышечного синапса. Пессимум Введенского можно получить и при прямом, но более частом раздражении мышцы (около 200 имп/с). Контрактура. Контрактурой называется состояние обратимого местного устойчивого сокращения. Оно отличается от тетануса отсутствием распространяющегося потенциала действия. При этом может наблюдаться длительная локальная деполяризация мышечной мембраны, например при калиевой контрактуре, или же мембранный потенциал, близкий к уровню покоя, в частности при кофеиновой контрактуре. Кофеин при нефизиологически высоких (миллимолярных) концентрациях проникает в мышечные волокна и, не вызывая возбуждения мембраны, способствует высвобождению Са2+ из саркоплазматического ретикулума; в результате развивается контрактура. При калиевой контрактуре степень стойкой деполяризации и сократительного напряжения волокна зависит от концентрации К+ в наружном растворе.
|
||||||||||
|
|
Последнее изменение этой страницы: 2016-08-14; просмотров: 1256; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.149.25.26 (0.013 с.) |