Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Органов нервной системы у человека.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Врожденные пороки развития, как правило, носят мультифакториальный характер. Они могут быть нарушением в генетическом аппарате больного или же возникать под действием тех или иных экзогенных или эндогенных факторов, обладающих тератогенным действием. Описано много форм врожденных пороков развития, среди которых моно назвать такие, как: 1) Дефекты смыкания нервной трубки (при неполном слиянии краев нервного желобка). Следствием этого может быть отсутствие головного мозга – анэнцефалия. 2) Удвоение и расщепление спинного мозга. 3) Полость или киста спинного мозга, заполненная жидкостью (сирингомиелия). 4) Нейродермальные свищи (сообщение между спинным мозгом и кожей), которые могут привести к развитию менингита. 5) Дефекты формирования срединной линии головного мозга могут вызвать недоразвитие промежуточного мозга и формирование единого желудочка переднего мозга. 6) Микроцефалия (окружность головы в два раза меньше нормы). 7) Макроцефалия как следствие генетически обусловленных нарушений метаболических процессов или кистозных образований паутинной оболочки мозга. 8) Гидроцефалия – расширение желудочков мозга и увеличение головы вследствие чрезмерного объема ликвора. 9) Аномалии развития мозжечка с полным отсутствием червя (сочетается с кистозным расширением IV желудочка). 10) Грыжевое выпячивание оболочек мозга (менингоцеле) или оболочек и вещества спинного мозга через дефект позвонков (обычно в поясничном отделе). 11) Ограничение подвижности спинного мозга, когда его концевые нити окружены фиброзной и жировой тканью. Это препятствует нормальному росту спинного мозга по мере роста ребенка. 12) Протрузия (выпячивание) ствола мозга и мозжечка в шейный отдел спинного мозга через большое затылочное отверстие. II. Филогенез органов чувств.
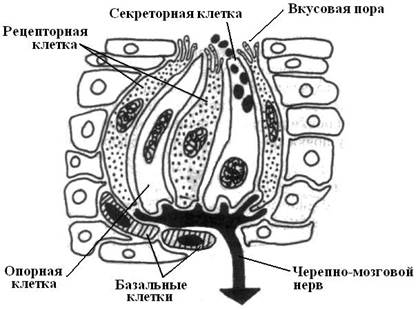
Под органами чувств, или сенсорными системами, понимают совокупность органов и структур, обеспечивающих восприятие различных раздражителей действующих на организм и поступающих как из внешней среды, так и внутренней, что необходимо для адаптации организма к конкретным условиям. Способность к элементарному анализу окружающей среды с самого начала эволюции животного мира базируется на таком фундаментальном свойстве всего живого как раздражимость, и совершенствовалась в ходе их приспособлений к изменяющимся условиям окружающей среды. Одноклеточные животные в ответ на механические, химические и электромагнитные раздражители отвечают таксисами – направленными движениями от источника раздражения (отрицательный таксис) или, наоборот, к нему (положительный таксис). Как показали эксперименты на амебах, в основе таксисов лежит воздействие раздражителей на рецепторные участки плазматической мембраны, что сопровождается включением АТФ-АТФазной системы, активизирующей аппарат управления движениями псевдоподий. В ходе эволюции многоклеточных организмов выделяются клетки, специализирующиеся на рецепторных функциях. При этом в случае механических воздействий рецепторные клетки реагируют на деформацию клеточной мембраны изменением проницаемости механозависимых ионных каналов, что приводит к изменению заряда мембраны и формированию градиента потенциала. Механорецепцией обладают: свободные нервные окончания, большая часть соматосенсорных рецепторов, органы боковой линии, рецепторы линейного и углового ускорения и органы слуха. В основе химической чувствительности лежит способность рецепторных молекул, расположенных с наружной стороны клеточной мембраны, распознавать различные вещества. Такие молекулы соединены с ионным каналом, закрытым при обычном состоянии, а при контакте со стимулирующим веществом этот канал открывается и изменяется локальный заряд мембраны. Химической чувствительностью обладают обонятельные и вомероназальные рецепторы, осморецепторы гипоталамуса, вкусовые рецепторы, кожные хеморецепторы костных рыб и др. Электромагнитная чувствительность построена на способности электромагнитных волн оказывать влияние на органические молекулы, находящиеся внутри специализированных клеток. На этом построена фоторецепция. Однако, одни фоторецепторы воспринимают электромагнитные волны в световом диапазоне (глаза, пинеальный комплекс), другие – в инфракрасном диапазоне (этмоидные органы термочувствительности у некоторых змей и ампулярные электрорецепторные органы рыб). Способностью реагировать на напряженность электромагнитного поля обладают утконос и, вероятно, дельфины. В самой центральной нервной системе позвоночных отсутствуют какие-либо специализированные рецепторы и поэтому операции на мозге безболезненны. Однако, есть данные о наличии специализированных нервных окончаний в отдельных парасимпатических узлах. В зависимости от расположения рецепторов в теле и характера воспринимаемых раздражений их подразделяют на: 1) экстероцепторы – реагируют на стимулы, поступающие из внешней среды (например, ухо, воспринимающее звуки, глаз, орган обоняния); 2) интероцепторы – воспринимают сигналы, поступающие из внутренней среды организма (например, рецепторы сонных артерий, реагирующие на изменение кровяного давления и содержание в крови СО2); 3) проприоцепторы – реагируют на сигналы, связанные с положением и движением частей тела и сокращением мышц. Все рецепторы преобразуют энергию стимула в локальный электрический сигнал. Последний возбуждает нервный импульс в сенсорном нейроне, который передается в мозг, где на него вырабатывается нужная ответная реакция. Все рецепторы преобразуют энергию стимула в локальный электрический сигнал. Последний возбуждает нервный импульс в сенсорном нейроне, который передается в мозг, где происходит анализ действующих раздражителей и вырабатывается на них нужная ответная реакция. В зависимости от общности физической природы раздражителей сенсорные системы классифицируют на: хеморецепторные, механорецепторные, фотрецепторные, терморецепторные, электрорецепторные, рецепторы магнитных полей. Эволюция рецепторов шла как в направлении их специализации в восприятии различных форм и видов раздражителей, так и в направлении совершенствования вспомогательных структур, усиливающих действие раздражителя, ставшего адекватным, и препятствующих проникновению других видов раздражителей. В зависимости от характера рецепторных элементов органы чувств делят на три типа. Первый тип – это органы чувств, у которых рецепторами являются специализированные нейросенсорные клетки (например, орган зрения орган обоняния), преобразующие внешнюю энергию в нервный импульс. Второй тип органов чувств имеет не нервные рецепторные клетки, а эпителиальные. От последних преобразованное раздражение передается чувствительным нейронам, которые воспринимают возбуждение сенсоэпителиальных клеток и порождают нервный импульс (например, у позвоночных органы слуха, равновесия, вкуса). К третьему типу относятся пропреопцептивная, кожная и висцеральная системы. Переферические рецепторные отделы в них представлены различными инкапсулированными и неинкапсулированными рецепторами, в состав которых обязательно входят ветвящиеся терминальные участки нервных волокон. Инкапсулированные рецепторы при всем их разнообразии всегда, кроме ветвления нервных волокон (осевого цилиндра), состоят из глиальных клеток. Снаружи они бывают покрыты слоистой соединительнотканной капсулой. К ним относятся, например, у млекопитающих осязательные тельца Мейснера, нервно-мышечные и нервно – сухожильные веретена и др. Неинкапсулированные нервные окончания состоят только из ветвлений осевого цилиндра. Они воспринимают холод, тепло и боль. Хеморецепторные системы. Одними из наиболее ранних форм чувствительности являются хеморецепторные системы, т.к. они затрагивают процессы обмена веществ, лежащие в основе всех проявлений жизни. Кроме того, эти системы служат средством выявления неблагоприятной среды обитания и лежат в основе способов коммуникации особей и их полового поведения. Развитие хеморецепции шло по двум главным направлениям: формирование контактной химической чувствительности (вкус) и дистантной (обоняние), что имеет важное значение для получения наиболее полной информации по признакам пригодности пищи. В основе хеморецепции одноклеточных животных лежит присутствие в их цитоплазматической мембране рецепторных молекул, способных избирательно реагировать на определенные химические вещества (например, на аминокислоты). У низших беспозвоночных хеморецепция осуществляется, видимо, за счет свободных нервных окончаний в форме терминальных ветвлений дендритов чувствительных нейронов, присутствующих в определенных участках тела и передающих информацию в нервные центры. Вкусовую избирательную активность проявляют уже гидры и актинии. В эксперименте они проглатывают съедобную добычу, а несъедобную отбрасывают. Избирательной хеморецепцией обладают и улитки, питающиеся водорослями. Об этом свидетельствуют эксперименты по кормлению их экстрактами различных водорослей. У ракообразных свободные хеморецепторы обнаружены в области антеннул, на ротовых органах и на внутренней поверхности клешней. У более высокоорганизованных беспозвоночных наряду со свободными нервными окончаниями присутствуют более сложно устроенные хеморецепторы дистантного действия. Это клеточные комплексы, в которых кроме чувствительных хеморецепторных клеток, присутствуют и вспомогательные клетки. Рецепторные имеют ветвящиеся чувствительные отростки, погруженные в жидкость и защищенные кутикулярным куполом с порами. Такого рода рецепторы располагаются на антеннах и имеют ведущее значение в поисках пищи, половых партнеров и передаче биологически значимой информации. Они реагируют на химические агенты, оказывающие привлекающее воздействие (аттрактанты) и вызывающие избегание (репелленты). На этом свойстве насекомых в сельском хозяйстве построено вырабатывание у пчел запаховых условных рефлексов, что способствует опылению кормовых овощей и фруктовых деревьев. Хеморецепция у насекомых лежит в основе строения и функции органов обоняния и вкуса. У общественных насекомых хеморецепция играет большую коммуникативную роль (опознавание своих и чужих, сигналы тревоги). Контактная хеморецепция насекомых осуществляется главным образом рецепторными органами ротового аппарата и лапок. Органы обоняния У позвоночных обонятельные клетки имеют отростки, погруженные в слизь, покрывающую поверхность органа обоняния. У круглоротых и рыб таким органом являются обонятельные ямки или подкожные мешки, расположенные рядом со ртом. У акуловых обонятельные ямки находятся на вентральной стороне рыла, а у костистых рыб – на дорсальной. У амфибий эти мешки открываются не только наружу, но и в ротовую полость. У рептилий в верхних дыхательных путях имеются зачатки обонятельных раковин. Дальнейшее развитие они получают у птиц и особенно у млекопитающих. Клеточный состав органов обоняния позвоночных такой же, как у беспозвоночных. У них собственно рецепторные клетки – это преобразованные нервные клетки, воспринимающие раздражения дендритами с пучком ресничек, погруженных в слой слизи. По аксонам обонятельных нейронов импульсы поступают к клеткам обонятельной луковицы (часть головного мозга). У высших млекопитающих (и у человека) от обонятельной луковицы возбуждение передается в подкорковые центры, а затем в кору больших полушарий в область крючка извилины морского коня. Обоняние играет важную роль в пищевом, защитном, половом поведении и других видах поведения животных. Запах выделений специальных желез у многих млекопитающих служит средством маркировки границ охотничьей территории, узнавания особей своего вида, другого пола и т.д.. Органы вкуса позвоночных образованы вкусовыми почками, которые состоят из нескольких десятков рецепторных клеток, окруженных поддерживающими клетками. На апикальном конце рецепторных клеток имеется пучок микроворсинок, сходящихся в области поры, куда проникает раздражитель. Рецепторные клетки оплетены дендритами чувствительных нейронов. У рыб вкусовые почки находятся не только в стенках ротовой полости, но и в глотке, жабрах, коже и усиках. У амфибий они сосредоточены только на языке, а у млекопитающих – на языке, небе и миндалинах в так называемых вкусовых сосочках. Возбуждение по чувствительным нейронам передается в таламус, а оттуда к корковым центрам в район нижнего конца центральной извилины больших полушарий.
Все разнообразие вкусовых ощущений возникает в результате их смешения и взаимодействия как с тактильными и болевыми рецепторами, так и с обонянием. Так, «едкий» вкус связан с раздражением болевых рецепторов полости рта, «острый» зависит от примеси обонятельных ощущений, «кисловатый» при вяжущем действии дубильных веществ тактильной модальности. Хеморецепторы внутренних органов. Значение хеморецепторов для органов пищеварения было показано И.П.Павловым. Например, хеморецепторы двенадцатиперстной кишки при раздражении соляной кислотой желудочного сока вызывают рефлекс замыкания пилорического сфинктера и усиление секреции поджелудочной железы. Имеются хеморецепторы и в верхних дыхательных путях позвоночных. В бронхах и альвеолах они реагируют на повышенную концентрацию CO2 и рефлекторно вызывают одышку. Обнаружены хеморецепторы в почечной паренхиме, мочевом пузыре, половых органах, скелетной мускулатуре и некоторых других органах. Механорецепторные системы. Механорецепция развилась как специализированная система, обеспечивающая получение информации о предметах внешнего мира и организации движений. В процессе эволюции механорецепторы по функциональному назначению разделились на: тактильные (осязательные), слуховые, статокинетические, проприоцептивные и интероцептивные. Характерной структурой обеспечивающей тактильную рецепцию беспозвоночных являются разбросанные в покровах тела первично чувствующие клетки. Их наружный конец снабжен плотным отростком (щетинкой, волоском), а базальный переходит в нервный отросток, передащий нервный импульс нейронам определенного нервного центра. Даже небольшое соприкосновение волоска с внешним предметом приводит к деформации мембраны, вызывая ее деполяризацию, что сопровождается потенциалом действия в виде нервного импульса.
У кольчатых червей присутствуют и более сложные кожные механорецепторы в виде группы чувствующих клеток, окруженных обкладочными эпителиальными клетками. У моллюсков тоже, кроме отдельных чувствующих клеток, имеются более сложно устроенные механорецепторные органы. У членистоногих имеются отдельные чувствующие клетки (или их группы) с отростками, входящими в подвижную или неподвижную, разнообразной формы, хитиновую щетинку. Располагаются такого рода рецепторы в различных участках тела. Так, у саранчи и чешуекрылых насекомых они располагаются на голове. Они приводятся в движение током воздуха и дают сигналы для корректировки полета. У мух соответствующе рецепторные органы находятся на антеннах.
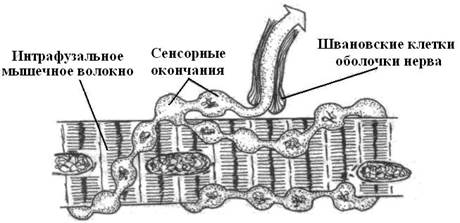
Наряду с особыми чувствующими клетками уже у низших червей имеются свободные нервные окончания, оканчивающиеся на клетках покровного эпителия. Такого рода механорецепторы имеются и у позвоночных животных. Это тактильные окончания. Благодаря ним животные реагируют на прикосновения. Они имеются у рыб, амфибий, рептилий, птиц и млекопитающих и располагаются в коже. У амфибий наиболее густо они усеивают околоротовую область и конечности. У пресмыкающихся механические раздражения воспринимаются осязательными волосками, расположенными на роговых чешуйках и связанными осязательными пятнами – скоплениями рецепторных клеток, лежащими под эпидермисом. У птиц и млекопитающих в связи с развитием перьевого и волосяного покрова они образуют сплетения вокруг основания пера или волосяных луковиц. У некоторых грызунов и хищных в качестве органа осязания чрезвычайного развития достигают так называемые вибриссы морды. Их сложный рецепторный аппарат дает разностороннюю информацию не только о прикосновении и давлении, но и о форме встречаемых предметов, окружающей обстановке. Исследования кожной чувствительности у приматов и человека показали более быстрые реакции на прикосновение к участкам кожи, покрытым волосами в сравнении с безволосыми. К тактильным рецепторам кожи у этих млекопитающих относятся также поверхностно расположенные диски Меркеля и тельца Мейснера, реагирующие на прикосновения, а также тельца Пачини, реагирующие на давление и вибрацию. Диски Меркеля это комплексы клеток с нервными терминалями, локализующиеся в базалее эпидермиса. У человека их особенно много в коже ладоней и подошв. Телец Мейснера – это инкапсулированные нервные окончания. Их особенно много в сосочковом слое кожи ладоней, подошв, в коже губ, век, наружных половых органов и сосков молочной железы. Тельца Пачини являются самыми крупными инкапсулированными нервными окончаниями, локализующимися глубоко в коже и подкожной клетчатке преимущественно пальцев, наружных половых органов и в молочной железе. Они воспринимают чувство давления. Имеются и другие инкапсулированные механорецепторы. Кинестезия. Кинестезией принято называть чувство положения и движения конечностей. Проприоцептивную информацию о взаиморасположении частей тела и состоянии двигательной системы у низших беспозвоночных дают механорецепторы находящиеся в основном в самих рганах движения и в местах тела, где они отходят от него. У членистоногих такого рода рецепторы формируются в области кутикулярных сочленений и растяжения мышц. У раков и омаров такими рецепторами растяжения снабжены разгибатели брюшка. В связи с усложнением движения у позвоночных развивается сложная система проприоцепции – мышечного и суставного чувства. Это чувство складывается из информации, поступающей от кожных, мышечных, суставных и сухожильных рецепторов, а также от рецепторов углового и линейного ускорения. Основную роль при этом играет информация, приходящая от мышц и сухожилий. Проприоцепция слагается как из этой информации, так и из сигналов, поступающих от суставов и суставных связок. Хотя информация от тех или иных проприорецепторов не осознается животными или человеком, однако общее состояние скелетно-мышечной системы дает вполне осознанное ощущение. Для точной координации движений и анализа положения тела возникли специализированные рецепторы. Так, у некоторых рыб в плавниковых мышцах имеются медленно адаптирующиеся механорецепторы, которые сигнализируют об их тоническом напряжении. Располагаются они не в самих мышечных волокнах, а в окружающей их соединительной ткани. У амфибий впервые появляются мышечные веретена. Они включают видоизмененные мышечные волокна, на которых располагается плоское или спиральное нервное окончание. Последнее посылает в мозг быстрые сигналы при удлинении мышцы.
У птиц и млекопитающих веретена состоят из нескольких мышечных волокон с первичными быстро реагирующими и вторичными медленно реагирующими гроздевидными рецепторами, формирующими сигналы как при сжатии, так и при растяжении мышцы. Для точной координации движений и анализа положения тела возникли разнообразные суставные и сухожильные рецепторы. В соединительнотканной капсуле суставов имеется много специализированных рецепторов, напоминающих немного модифицированные кожные рецепторы типа телец Руффини, и Пачини. Кроме того, в суставных связках присутствуют немиелинизированные свободные нервные окончания. В сухожилиях имеются миелинизированные рецепторы Гольджи с медленной адаптацией к растяжению-сжатию.
Обильная иннервация суставов характерна только для млекопитающих и птиц. У низших позвоночных как сами суставные сумки, так и связки практически не иннервированы, а суставные рецепторы расположены в окружающей соединительной ткани. Органы боковой линии. У рыб и живущих в воде амфибий присутствует особый вид механорецепторов – органы боковой линии. Они представляют собой отдельные ямки или систему каналов, которые идут вдоль боковой поверхности тела в виде полоски. И каналы и ямки сообщаются с внешней средой порами. В каналах и на дне ямок расположены чувствительные бугорки – нейромасты. В некоторых случаях (например у шпорцевой лягушки) последние лежат прямо на поверхности тела. В нейромастах имеются волосковые клетки, относящиеся к безотростковым механорецепторам. Волоски этих клеток погружены в студенистые бугорки, которые называются купулами. На поверхности рецепторных клеток имеются два ряда ресничек, одна из которых неподвижная – киноцилия, а другая тонкая подвижная – стереоцилия. Если стереоцилии отклоняются в сторону киноцилий, волосковая клетка деполяризуется, если же стереоцилии отклоняются в противоположную от киноцилий сторону, то волосковая клетка гиперполяризуется. В нейромастах волосковые клетки располагаются попарно, чем достигается различение направленности движения воды. К каждой волосковой клетке подходят эфферентное и афферентное нервные волокна, чем обеспечивается дискретность информации. Органы равновесия. Механорецепция как средство ориентации в пространстве наблюдается уже у простейших. Инфузории различают «верх» и «низ», используя твердые внутриклеточные включения как прототипы статоцистов высших животных. У медуз по краю колокола располагаются статоцисты – замкнутые пузырьки, в полости которых статолиты, состоящие из CaSO4, своей тяжестью раздражают первичные рецепторные клетки, сигнализируя о положении тела в пространстве. У ракообразных в статоцистах, расположенных у основания антеннул, образуются вертикальный и горизонтальный каналы, движение жидкости в которых раздражает волоски групп клеток, осуществляющих рецепцию углового ускорения. Роль статолитов в них играют песчинки. У позвоночных основную роль в определении положения и движения тела играет вестибулярный аппарат. Он развивается на основе органа боковой линии рыб, из полости которого выпячиваются мешочки, дающие начало полукружным каналам. У миксин образуется единственный вертикальный канал, у миног их два, а у остальных их три. Они дают информацию о движениях в любом направлении трехмерного пространства. Вестибулярный аппарат высших позвоночных животных представлен двумя мешочками и тремя полукружными каналами с ампулами. Один из мешочков круглый (саккулюс), другой – овальный (утрикулюс). В них имеются участки с сенсорными (чувствительными) волосковыми клетками, которые входят в состав, так называемых макул. В последних присутствуют еще и опорные клетки. Поверх таких скоплений клеток располагается студенистая отолитовая мембрана с вкрапленными в нее кристаллами карбоната кальция – отолитами. На верхней поверхности сенсорных клеток имеется кутикула, от которой отходят неподвижные чувствительные волоски – стереоцилии и одна подвижная ресничка – киноцилия, располагающаяся полярно относительно пучка стереоцилий. Стимуляция сенсорных клеток обеспечивается скольжением по их поверхности отолитовой мембраны. Возникающее при этом возбуждение воспринимается чувствительными нейронами, дендриты которых образуют синапсы на сенсорных клетках. Возникающий при этом нервный импульс передается в соответствующую часть вестибулярного анализатора.
Макулы овальных мешочков – это места восприятия линейных ускорений, рецепторы гравитации (земного притяжения), связанные с изменением тонуса мышц, определяющих установку тела. Макулы круглых мешочков, кроме гравитации, воспринимают и вибрационные колебания. Рецепторные образования вестибулярного аппарата присутствуют и в ампулах полукружных каналов (рис. 19).
Они воспринимают сигналы главным образом о движении головы в трех взаимоперпендикулярных направлениях и называются кристами (ампулярными гребнями). Кристы имеют вид поперечных складок, состоящих из волосковых и поддерживающих эпителиальных клеток, над вершинами которых располагается желеобразный купол. Последний смещается при движении эндолимфы в полукружных каналах и раздражает волосковые клетки. Их возбуждение воспринимается волокнами вестибулярного нерва и передается в соответствующие участки головного мозга, откуда нервные импульсы поступают скелетным мышцам.
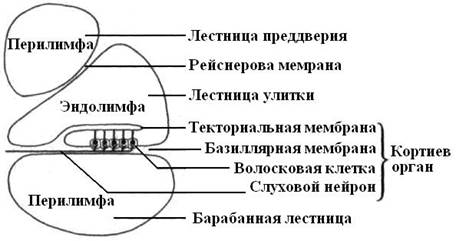
Орган слуха. Приспособление механорецепторов к восприятию звуков в ходе эволюции было связано не только с появлением специфических звуковоспринимающих клеток, но и вспомогательных структур, позволяющих дифференцировано реагировать на звуки. В этом отношении животные обнаруживают большое разнообразие, связанное с их экологией. Среди беспозвоночных органы слуха имеются у членистоногих. Они служат для них органами коммуникации, а в ряде случаев позволяют спасаться от врагов. У одних органы слуха представлены тимпанальными мембранами, у других – хордотональными слуховыми органами. Тимпанальные мембраны натянуты на прочной хитиновой рамке и выполняют роль барабанной перепонки. Под мембраной лежит трахейный пузырь, а между ним и мембраной – группы рецепторных клеток. Хордотимпальные слуховые органы состоят из струнообразной связки, натянутой над покровами тела, и сенсорных клеток, которые одним концом соединяются со связкой, а другим – с покровами. Связка колеблется в унисон звуковым волнам, поступающим из окружающей среды, и возбуждает рецепторные клетки. Орган слуха позвоночных развивается вместе с органом равновесия из утолщения эктодермы в задней части головы. Этот зачаток углубляется и принимает вид пузырька, который в дальнейшем разделяется на 2 отдела. Из верхнего отдела образуются полукружные каналы и овальный мешочек, а из нижнего формируется полое выпячивание – лагена и круглый мешочек. Все эти образования заполнены жидкостью – эндолимфой и составляют так называемый перепончатый лабиринт. Он окружен стенками скелета, а промежуток между ними и перепончатым лабиринтом заполнен перилимфой. У круглоротых, рыб и безногих амфибий со слуховой рецепцией связаны 2 сосочка круглого мешочка. Для амфибий характерен амфибийный сосочек, а для остальных тетрапод – базилярный сосочек, в которых звуковоспринимающую функцию выполняют рецепторные волосковые клетки. У рептилий, птиц и однопроходных имеется вытянутый слепой вырост – лагена. У птиц она к тому же изгибается, а у млекопитающих многократно закручивается, образуя улитку. У млекопитающих литка разделяется тремя перепонками на 3 части. Две из них – это лестница преддверия и барабанная лестница, содержат перилимфу и соединяются между собой на верхушке улитки через небольшое отверстие. Средний канал улитки (перепончатый лабиринт) содержит эндолимфу. Он отделен от барабанной лестницы базилярной мембраной, а от лестницы преддверия – рейснеровой. Базиллярная мембрана состоит из поперечно ориентированных коллагеновых волокон («струн»), длина которых по направлению к основанию улитки постепенно уменьшается. Эти волокна резонируют в унисон звукам различной высоты.
Располагающийся на базилярной мембране звуковоспринимающий кортиев орган состоит из сенсорных волосковых и поддерживающих эпителиальных клеток, которые подразделяются на наружные и внутренние.
На апикальных участках сенсорных клеток имеются пучки коротких уплотненных микроворсинок, способных отклоняться – это стереоцилии. Стереоцилии наружных сенсорных клеток своими вершинами прикасаются к поверхности лежащей над ними текториальной мембраны. Наружные сенсорные эпителиоциты значительно чувствительнее к звукам большей интенсивности, чем внутренние.
Первичноводные позвоночные довольно широко используют коммуникации, связанные с испусканием звуков. Очень часто они для этого используют растяжение плавательного пузыря. Звуковоспринимающую функцию у них выполняют рецепторные клетки макул круглого мешочка и лагены. Звуковые волны эффективно передаются в воде, поэтому у первичноводных животных нет специализированных образований для фильтра и усиления звуков. Эту функцию выполняет все тело, по тканям которого колебания воды передаются практически без потерь. У наземных позвоночных условия звукопередачи на внутреннее ухо изменяются, т.к. воздействие звуковой волны оказывается слишком слабым для рецепторного аппарата внутреннего уха. Поэтому у них в эволюции сформировались специальные приспособления для усиления звуковых сигналов – среднее и наружное ухо. Все особенности структур среднего уха направлены на усиление внешних звуковых сигналов. Вся система среднего уха сложилась на основе морфогенетических изменений одной из висцеральных щелей – брызгальца, наружная часть которого несколько расширилась, образовав барабанную полость, и закрылась барабанной перепонкой, а внутренняя сузилась и превратилась в евстахиеву трубу – узкий канал, конец которого, как и брызгальца открывается в глотку. В среднем ухе имеется набор косточек, которые передают колебания от барабанной перепонки к овальному окну. У амфибий, рептилий и птиц сигналы передаются через столбик (колумелла), а у млекопитающих – через наковальню, молоточек и стремечко.
У змей и некоторых других рептилий барабанная перепонка редуцирована. У них колебания воздуха передаются от квадратной черепа через короткую связку колумелле, а от нее к овальному окну. Наружное ухо присуще птицам и млекопитающим. Птицы обладают слухом в большом частотном диапазоне, что обеспечивается организацией рецепторного аппарата и особенностями наружного уха. У птиц лагена в сравнениями с рептилиями увеличена в размерах, но еще не формирует завитков, как у млекопитающих. Слуховые рецепторные клетки у птиц располагаются в лагене в перепончатом лабиринте, который во многом похож на лабиринт млекопитающих. У млекопитающих слуховой аппарат внутреннего уха представляет собой длинный рукав, который закручен в виде улитки. Исключение составляют однопроходные (яйцекладущие), у которых практически прямой рукав внутреннего уха (нет улитки). Полость среднего уха соединяется с ротовой полостью евстахиевой трубой и отличается главным образом большой сложностью передаточного аппарата, состоящего из трех слуховых косточек. Особая мышца, прикрепляющаяся к рукоятке молоточка, натягивает барабанную перепонку, а другая – мышца стремени, наклоняет стремечко и натягивает перепонку овального окна. Рефлекторно сокращением этих мышщ уменьшается амплитуда колебаний всего аппарата при более сильных звуках. У млекопитающих значительного развития достигает наружный слуховой проход и связанные с ним наружные ушные раковины. Последние собирают звуковые волны и направляют их в проход. Ушная раковина имеет хрящевой скелет и приводится в движение особой мускулатурой. У китообразных в связи с их образом жизни ушная раковина отсутствует, наружный слуховой проход редуцирован, и звуки проводятся к среднему и внутреннему уху через жировые и костные ткани черепа. Одной из основных характеристик слуха является восприятия звуков определенного диапазона частот. Наибольшее разнообразие в этом отношении характерно для позвоночных животных. Так, рыбы чутко реагируют на инфразвуковые и низкочастотные колебания, издаваемые плавающими в воде животными. Сами рыбы издают с аналогичным диапазоном звуки типа: хрюканья, крика, тарахтенья, гудков, скрежета и др. Пресмыкающиеся слышат звуки более высокого тона, чем амфибии. Большинство птиц наиболее чувствительны к звукам 3,0 – 3,5 кГц, собаки – 35 кГц, летучие мыши и дельфины до 100 кГц, человек от 16 до 18 кГц. В то же время тимпанальные органы кузнечиков слышат звуки около 200 кГц. Согласованная деятельность обоих ушей обеспечивает по разности силы и фазовому сдвигу звуковых колебаний, приходящих в одно и другое ухо, определять направление к источнику звука.
|
||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-06-26; просмотров: 483; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.14.247.170 (0.013 с.) |