
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Типологическая концепция вида. Главные концепции видаСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
12. Впервые термин и концепцию генофонда сформулировал русский советский генетик А. С. Серебровский в 1928 году[:
Четвериков выдвинул предположение о насыщении видов в природе возникающими мутациями и подчеркнул значение генетических процессов (мутация, свободное скрещивание, естественный отбор) и изоляции в видообразовании и эволюции, тем самым связав теорию эволюции Дарвина и генетики. Заложил основы эволюционной генетики. В работах 20-х гг. С. С. Четвериков обосновывает три основные посылки популяционной генетики: Мутационный процесс в природных условиях протекает точно так же, как и в условиях лаборатории. Поэтому мы вправе распространять по крайней мере некоторые выводы, полученные в лаборатории, на природные ситуации. Один из таких выводов — непрерывное во времени возникновение новых мутаций у всех видов живых организмов, другой — рецессивность большинства вновь появляющихся мутаций по отношению к аллелям дикого типа, распространенным в природных популяциях. Характернейшей чертой природных популяций является преобладание в них панмиксии, что делает возможным приложение Закон Харди — Вайнберга. Исходя из признания важности для понимания эволюции проблем природы изменчивости и ее поддержания в популяциях Четвериков анализирует процесс возникновеия мутаций в природе, их поведение в условиях свободного скрещивания и роль отбора в их распространении. Рассматривая популяцию как элементарную структуру эволюции, он вводит понятия о непрерывном мутационном давлении, о генофонде и о генотипической среде, он предсказывает насыщенность вида гетерозиготными мутациями (накопленный резерв наследственной изменчивости) и возможность их выявления, выход в гомозиготное состояние при инбридинге, а также неселективные различия между генофондами популяций в условиях изоляции. В этой статье Четвериков показал совместимость принципов генетики с теорией естественного отбора, заложил основы эволюционной генетики. Его идеи и предсказания блестяще подтвердили в опытах на дрозофиле его ученики С. М. Гершензон, Д. Д. Ромашов, Е. И. Балкашина и Н. В. Тимофеев-Ресовский. Последний, наряду с Ф. Добржанским, привнес в европейскую и американскую генетику разработанную Четвериковым методологию изучения популяций и микроэволюции.
13. Закон Харди — Вайнберга — это закон популяционной генетики — в популяции бесконечно большого размера, в которой не действует отбор, не идет мутационный процесс, отсутствует обмен особями с другими популяциями, не происходит дрейф генов, все скрещивания случайны — частоты генотипов по какому-либо гену (в случае если в популяции есть два аллеля этого гена) будут поддерживаться постоянными из поколения в поколение и соответствовать уравнению:
Где
14. Дрейф ге́нов или гене́тико-автомати́ческие проце́ссы — явление ненаправленного изменения частот аллельных вариантов генов в популяции, обусловленноеслучайными статистическими причинами.Один из механизмов дрейфа генов заключается в следующем. В процессе размножения в популяции образуется большое число половых клеток — гамет. Большая часть этих гамет не формирует зигот. Тогда новое поколение в популяции формируется из выборки гамет, которым удалось образовать зиготы. При этом возможно смещение частот аллелей относительно предыдущего поколения.Первые работы по изучению случайных процессов в популяциях были проведены в начале 1930-х годов Сьюэлом Райтом в США, Роналдом Фишером в Англии, а также В. В. Лисовским, М. А. Кузнецовым, Н. П. Дубининым и Д. Д. Ромашовым в СССР. Понятие «дрейф генов» было введено в оборот Райтом (1931), асинонимичноепонятие «генетико-автоматические процессы в популяциях»— Дубининым и Ромашовым (1932).В ходе эволюции популяций дрейф генов взаимодействует с другими факторами эволюции, прежде всего с естественным отбором. Соотношение вкладов этих двух факторов зависит как от интенсивности отбора, так и от численности популяций. При высокой интенсивности отбора и высокой численности популяций влияние случайных процессов на динамику частот генов в популяциях становится пренебрежимо малым. Наоборот, в малых популяциях при небольших различиях по приспособленности между генотипами дрейф генов приобретает решающее значение. В таких ситуациях менее адаптивный аллель может зафиксироваться в популяции, а более адаптивный может быть утрачен. Конечным результатом дрейфа генов является полное устранение одного аллеля из популяции и закрепление (фиксация) в ней другого аллеля. Чем чаще тот или иной аллель встречается в популяции, тем выше вероятность его фиксации вследствие дрейфа генов. Расчеты показывают, что вероятность фиксации нейтрального аллеля равна его частоте в популяции.
Н. Вавилов - автор концепции линнеевского вида как системы (1930). Инициатор создания многих научно-исследовательских учреждений. Член ЦИК СССР (1926-1935), президент Всесоюзного географического общества (1931-1940). Премия имени Владимира Ильича Ленина (1926). 6 августа 1940 арестован, обвинен в контрреволюционной вредительской деятельности и в июле 1941 года приговорен к расстрелу, замененному в 1942 году 20-летним заключением. Умер в больнице саратовской тюрьмы, посмертно реабилитирован в 1955 году. Применение нового закона позволило Вавилову поставить вопрос и о том, что все культурные растения Земли произошли из нескольких генетических центров. В 1924 году Вавилов организует экспедицию в Афганистан. Здесь он собирает исключительный по ценности материал. В 1926 году Вавилов совершает большую поездку по странам Европы, а также Северной Африки. В последующие годы Вавилов посетил Японию, Китай, а также Южную Америку. Он собрал столько образцов различных растений, что его теория получила полное подтверждение. Сразу после поездки выходит в свет его вторая важнейшая работа "Центры происхождения культурных растений". В 1920-1940 гг. Вавилов организовал экспедиции почти на все континенты земного шара, под его руководством была создана коллекция культурных растений, насчитывающая более 300 тыс. образцов и ставшая основой для выведения новых сортов. Опираясь на обширную селекционную деятельность, Вавилов создал учение о наследственной изменчивости, центральным звеном которого является сформулированный им закон гомологических рядов. Этот закон устанавливает, что близкие виды благодаря большому сходству их генотипов обладают сходной потенциальной наследственной изменчивостью, по мере усиления генотипических различий параллелизм наследственной изменчивости становится менее полным. Закон гомологических рядов отражает всеобщее и фундаментальное явление в живой природе. Пользуясь этим законом, по ряду морфологических признаков и свойств одного вида или рода организмов можно предвидеть существование соответствующих форм у другого вида или рода. Закон облегчает целенаправленный поиск нужных признаков, а тем самым и выведение новых видов
15. Миграция животных — регулярное передвижение популяции животных, в ходе которого особи из одной области обитания перемещаются в другую, но затем возвращаются обратно. Миграции наиболее распространены у птиц (перелеты птиц) и рыб (например, миграции тихоокеанского лосося). У зверей миграции изучены меньше, так как они ведут более скрытный образ жизни. Примеры миграций у животных — миграция северных оленей из тундры в лесотундру с наступлением зимы, она вызвана нехваткой и трудностями добычи корма в занесенной снегом тундре. Миграции животных имеют ярко выраженный приспособительный (адаптивный) характер и возникли в процессе эволюции у самых разных видов. Миграции можно разделить на активные (животные передвигаются сами) и пассивные (животные перемещаются на льдах или несутся водой). Активные миграции можно разделить по временным интервалам, в которые они происходят:
·Активные ·Сезонная миграция ·Суточная миграция ·Периодическая миграция
В настоящее время проблема вида переживает сильнейший кризис: с одной стороны появляются все новые и новые концепции, отражающие тот или иной аспект вида, тогда как "идеальной" концепции вида - во всей его целостности - никак не удается достигнуть; с другой стороны, биологи, раздраженные погоней за ускользающим призраком, призывают прекратить бесполезные споры по этому вопросу или вообще "избавиться от видов", а также упразднить все ранги (включая видовой) в биологической системе организмов и "просто" выделять монофилетические группы.Почти полтора века после Дарвина систематики и эволюционисты спорили о том, что такое вид, а в конце концов вернулись на тот же круг времени, когда Дарвин писал: "мы можем смутно предвидеть глубокий переворот в области естествознания; … мы будем относиться к видам таким же образом, как относятся к родам те натуралисты, которые допускают, что роды только искусственные сочетания, придуманные ради удобства; может быть, это не очень утешительная перспектива, но зато мы навсегда освободимся от тщетных поисков за неуловленной до сих пор и неуловимой сущностью слова "вид".Перспектива, действительно, не очень утешительная, тем более, что предвиденный Дарвином «глубокий переворот» в области естествознания, наконец, наступил и грозит уничтожить фактически все прошлые великие достижения в области систематики, в том числе иерархию (систему рангов) Линнея. Биологи поняли, что проблема вида - это частный случай «проблемы таксона», но вывели из этого неверное заключение о необходимости 'безранговой' классификации: " видовой ранг должен исчезнуть вместе со всеми другими рангами ".Очевидно, что наболевшая проблема вида грозит взорвать всю таксономическую теорию, хотя сама 'проблема вида' чрезвычайно узка в философском плане и решаема только в более широких систематических рамках. Некоторые исследователи полагали, что «общая теория вида, или эйдология, должна рассматривать вид всесторонне, в том числе и такие его черты, которые недоступны или не нужны для систематики», и что «ни старая, ни новая систематика не занимались и не призваны заниматься изучением внутривидовых отношений» [9], которые характеризовались как «сущность» вида.
16.Неродственное скрещивание, или аутбридинг, осуществляется между особями одной породы или между особями разных пород животных. При строгом отборе приводит к поддержанию свойств или улучшению их в ряду следующих поколений гибридов, т.к. в потомстве может получиться удачная комбинация генов, обеспечивающая формирование целого ряда хозяйственно-важных признаков. Близкородственное скрещивание, или инбридинг, проводится между братьями и сестрами или родителями и потомством. Этот тип скрещивания применяют в тех случаях, когда желают перевести большинство генов породы в гомозиготное состояние, т.е. для получения чистых линий, сохранения хозяйственно-важных признаков, повышения стабильности этих признаков для последующего скрещивания и получения эффекта гетерозиса.
Межлинейное скрещивание проводится между представителями чистых гомозиготных линий для того, чтобы избежать неблагоприятного действия рецессивных генов, перевести их в гетерозиготное состояние и вызвать эффект гетерозиса. Обычно для скрещивания используются представители нескольких линий. Отдаленная гибридизация, т.е. межвидовое скрещивание, известно у животных с древнейших времен. Чаще всего межвидовые гибриды стерильны, т.к. у них нарушается мейоз, что приводит к нарушению гаметогенеза. С глубокой древности человек использует гибрид кобылицы с ослом – мула, который отличается выносливостью и большой продолжительностью жизни. Преодоление бесплодия межвидовых гибридов животных – важная задача селекции. Кооперация, утверждал Кропоткин, хорошо видна в мире животных. Лошади и олени объединяются для совместной защиты от врагов, волки и львы собираются для охоты, пчёлы и муравьи сотрудничают самым разнообразным образом. […] Кропоткин к тому же утверждал, что люди расположены помогать друг другу без принуждения. Не нужно централизованной власти для того, чтобы подавать пример или заставлять людей поступать справедливо. Люди поступали так ещё до возникновения государства. В действительности же Кропоткин придерживался взгляда, что именно власть подавляет нашу природную склонность к сотрудничеству. Его приверженность началам народоправства и привела в итоге Кропоткина в царские застенки. Кропоткин писал о временных средневековых гильдиях - совместных, возникающих «по случаю» объединениях, создаваемых союзом единомышленников с общей целью и общим местом пребывания. Эти объединения можно было отыскать на судах, на строительстве больших общественных сооружений вроде соборов и повсюду, где «рыбаки, охотники, странствующие купцы, строители или оседлые ремесленники собирались вместе для занятия общим делом». После выхода в море капитан судна собирал на палубе команду и пассажиров, говоря им, что теперь они здесь одно целое и что успех плавания зависит от их совместных действий. Затем все избирали «судью» и «приставов» для взимания «пошлины» с нарушителей заведённых порядков. По завершении плавания весь сбор шел беднякам города, куда приставал корабль». 17. Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Некоторые этапы экспрессии генов могут регулироваться: это транскрипция, трансляция, сплайсинг РНК и стадия посттрансляционных модификаций белков.Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Экспрессия генов является субстратом для эволюционных изменений, так как контроль над временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме. СТРУКТУРА ВИДА, структура (инфраструктура) вида состоит из след. единиц: 1) полувид — географич. или экологич. раса, к-рая почти достигла состояния “молодого вида”, характеризующаяся достаточной морфофизиологич., географич., экологич., а в ряде случаев и репродуктивной особенностями; 2) подвиды — территориально разобщенные географич. расы, приспособленные к определенному местообитанию и отличающиеся морфофизиологич. признаками (напр., ель обыкновенная образует европейскую, финскую и сибирскую расы, или подвиды; обыкновенная белка на территории СССР имеет более 20 подвидов, отличающихся окраской, размерами и качеством меха); 3) экотипы — экологич. расы, приспособленные к тем или иным условиям и имеющие свои морфофизиологич. признаки (напр., выделяют лесную и парковую расу черного дрозда, экотипы растений кислых и щелочных почв и т. п.); 4) популяция — относительно обособленная на определенной территории группа свободноскрещивающихся особей, способная длительное время существовать, воспроизводиться, эволюционировать; популяции вида возникают в результате естественного отбора. Каждая популяция также имеет свою внутреннюю структуру. 18. Транскри́пция — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.Транскрипция катализируется ферментом ДНК-зависимой РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5'- к 3'- концу, то есть по матричной цепи ДНК РНК-полимераза движется в направлении 3'->5' Транскрипция состоит из стадий инициации, элонгации и терминации. Единицей транскрипции является транскриптон, фрагмент молекулы ДНК, состоящий из промотора, транскрибируемой части и терминатора. Дойдя до конца оперона, фермент встречает сигнал (в виде определенной последовательности нуклеотидов), означающий конец считывания. Готовая и- РНК отходит от ДНК и направляется к месту синтеза белков. В описанном процессе транскрипции можно выделить четыре стадии: 1) Связывание РНК-полимеразы с промотором; 2) Инициация - начало синтеза. Она заключается в образовании первой фосфодиэфирной связи между АТФ или ГТФ и вторым нуклеотидом синтезирующейся молекулы и-РНК; 3) элонгация - рост цепи РНК, т. е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные нуклеотиды в транскрибируемой нити ДНК. Скорость элонгации достигает 50 нуклеотидов в секунду; 4) терминация - завершение синтеза и-РНК. Интродуцированный, или чужеродный вид — некоренной, несвойственный для данной территории, преднамеренно или случайно завезённый на новое место в результате человеческой деятельности. Процесс освоения интродуцированного вида на новом месте (адаптации к новым экологическим условиям) называется акклиматизацией. Часто интродуцированные виды способны существенно изменить сложившуюся экосистему региона и стать причиной значительного сокращения или даже вымирания отдельных видов местной флоры и фауны. Такую интродукцию часто называют биологическим загрязнением. 19. ТРАНСЛЯЦИЯ, синтез белков (полипептидов) на рибосомах с использованием в качестве матрицы информационной рибонуклеиновой кислоты (и-РНК); завершающий этап реализации генетической информации в живых клетках. В ходе трансляции информация, записанная в нуклеиновых кислотах в виде генетического кода, переводится в последовательность аминокислот в синтезируемых белках. При этом четырёхбуквенныйнуклеотидный «язык» передаётся (лат. «трансляцио» – передача) «языком» двадцатибуквенным аминокислотным. Процесс трансляции разделяют на: инициацию — узнавание рибосомой стартового кодона и начало синтеза. элонгацию — собственно синтез белка. терминацию — узнавание терминирующего кодона (стоп-кодона) и отделение продукта. Рамка считывания Поскольку каждый кодон содержит три нуклеотида, один и тот же генетический текст можно прочитать тремя разными способами (начиная с первого, второго и третьего нуклеотидов), то есть в трех разных рамках считывания. За некоторыми интересными исключениями, значимой является информация, закодированная только в одной рамке считывания. По этой причине крайне важным для синтеза белка рибосомой является её правильное позиционирование на стартовом АУГ-кодоне — инициация трансляции. Критерии вида – это признаки, по которым сравнивают 2 организма, чтобы определить, относятся они к одному виду или к разным. ·Морфологический – внутреннее и внешнее строение. ·Физиолого-биохимический – как работают органы и клетки. ·Поведенческий – поведение, особенно в момент размножения. ·Экологический – совокупность факторов внешней среды, необходимых для жизни вида (температура, влажность, пища, конкуренты и т.п.) ·Географический – ареал (область распространения), т.е. территория, на которой живет данный вид. ·Генетико-репродуктивный – одинаковое количество и строение хромосом, что позволяет организмам давать плодовитое потомство. Критерии вида относительны, т.е. по одному критерию нельзя судить о виде. Например, существуют виды-двойники (у малярийного комара, у крыс и т.д.). Они морфологически друг от друга не отличаются, но имеют разное количество хромосом и поэтому не дают потомства. (То есть морфологический критерий не работает [относителен], но работает генетико-репродуктивный 20. Но́рма реа́кции — способность генотипа формировать в онтогенезе, в зависимости от условий среды, разные фенотипы. Она характеризует долю участия среды в реализации признака и определяет модификационную изменчивость вида. Чем шире норма реакции, тем больше влияние среды и тем меньше влияние генотипа в онтогенезе. Один и тот же ген в разных условиях среды может реализоваться в несколько проявлений признака (фенов). В каждом конкретном онтогенезе из спектра проявлений признака реализуется только один. Аналогично один и тот же генотип в разных условиях среды может реализоваться в целый спектр потенциально возможных фенотипов, но в каждом конкретном онтогенезе реализуется только один фенотип. Под наследственной нормой реакции понимают максимально возможную ширину этого спектра: чем он шире, тем шире норма реакции. Фенотипическое значение любого количественного признака (Ф) определяется, с одной стороны, его генотипическим значением (Г), с другой стороны — влиянием среды (С): Конце́пции ви́да — различные системы взглядов на понятие вид в биологии. Вид, как таксон, является базовой структурной единицей любой системы органического мира, от определения границ которого зависит структура всей таксономической иерархии. Понятие концепции вида тесно связано с другими направлениями, понятиями и предметами изучения биологии: критерии вида, видообразование, таксономия,биологическая систематика и другими. При этом проблема вида, ввиду наличия у этого таксона ряда уникальных свойств, может рассматриваться как самостоятельная область биологической науки.
21. Морфоз — ненаследственное изменение фенотипа организма в онтогенезе под влиянием экстремальных факторов среды. Морфозы имеют неадаптивный и часто необратимый характер. Часто это грубые изменения фенотипа, выходящие за пределы нормы реакции, в итоге развивается патология и может наблюдаться даже гибель организма.
Биологическая концепция вида - концепция, по которой основным критерием вида есть нескрещиваемость (репродуктивная изоляция). Формирование биологической концепции началось после 1750 года. Окончательно ее сформировал К. Иордан (1905) и Ф. Добжанский (1937). Эта концепция отличается от прежних тем, что учитывает чисто биологические постулаты. ·Виды определяются не различием, а особенностями. ·Виды, как правило, состоят не из особей, а из популяций. ·Решающий критерий вида - не плодовитость при скрещивании, а репродуктивная изоляция. ·Члены вида образуют генетическое единство, обладающее генофондом. ·Члены вида образуют репродуктивное сообщество: самцы и самки одного вида воспринимают друг друга как потенциальных партнеров для воспроизведения потомства. ·Вид - это экологическое единство: члены популяции взаимодействуют как одно целое с другими популяциями экосистемы. · 1)Что такое «Ароморфоз». Ароморфоз Внутривидовая конкуренция Если конкуренты принадлежат к одному виду, то взаимоотношения между ними называют внутривидовой конкуренцией. Конкуренция между особями одного вида является наиболее острой и жесткой в природе, поскольку они имеют одинаковые потребности в экологических факторах. Внутривидовую конкуренцию можно наблюдать в колониях пингвинов, где происходит борьба за жизненное пространство. Каждая особь удерживает свой участок территории и агрессивно настроена по отношению к соседям. Это приводит к четкому разделению территории внутри популяции. Внутривидовая конкуренция на том или ином этапе существования вида встречается почти всегда, поэтому в процессе эволюции у организмов выработались приспособления, снижающие ее интенсивность. Наиболее важные из них — способность к расселению потомков и охрана границ индивидуального участка (территориальность), когда животное защищает место своего гнездовья или определенный участок. Так, в период размножения птиц самец охраняет определенную территорию, на которую, кроме своей самки, не допускает ни одной особи своего вида. Такую же картину можно наблюдать и у некоторых рыб. Межвидовая конкуренция Если конкурирующие особи относятся к разным видам, то это межвидовая конкуренция. Объектом конкуренции может служить любой ресурс, запасы которого в данной среде недостаточны: ограниченная территория распространения, пища, участок для гнезда, элементы питания для растений. Результатом конкуренции может быть расширение области распространения одного вида за счет сокращения численности или исчезновения другого. В качестве примера можно привести активное расширение с конца XIX в. ареала длиннопалого рака, который постепенно захватил весь Волжский бассейн и добрался до Беларуси и Прибалтики. Здесь он начал вытеснять родственный вид — рака широкопалого. Конкуренция может носить достаточно острый характер, например в борьбе за гнездовую территорию. Такой тип называется прямой конкуренцией. В большинстве случаев эти конфликты происходят между особями одного вида. Однако часто конкурентная борьба протекает внешне бескровно. Например, на многих хищных животных, конкурирующих за пищу, другие хищники влияют не прямо, а опосредованно, через уменьшение количества пищи. То же происходит в мире растений, где при конкуренции одни влияют на другие опосредованно, через перехват питательных веществ, солнца или влаги. Этот тип носит название косвенной конкуренции. Конкуренция является одной из причин того, что два вида, слабо различающихся спецификой питания, поведения, образа жизни и т.д., редко сожительствуют в одном сообществе. Исследования причин и следствий межвидовой конкуренции привели к установлению особых закономерностей в функционировании отдельных популяций. Некоторые такие закономерности возведены в ранг законов. Исследуя рост и конкурентные взаимоотношения двух видов реснитчатых инфузорий, советский биолог Г.Ф. Гаузе провел ряд экспериментов, результаты которых опубликовал в 1934 г. Инфузории двух видов — Paramecium caudatum и Paramecium aurelia хорошо росли в монокультуре. Пищей им служили бактериальные или дрожжевые клетки, растущие на регулярно добавляемой овсяной муке. Когда Гаузе помещал оба вида в один сосуд, каждый вид сначала быстро увеличивал свою численность, но со временем P. aurelia начинала расти за счет P. caudatum, пока второй вид полностью не исчезал из культуры. Период исчезновения длился около 20 дней. Таким образом, Г.Ф. Гаузе сформулировал закон (принцип) конкурентного исключения, который гласит: два вида не могут существовать в одном местообитании (в одной и той же местности), если их экологические потребности идентичны. Поэтому любые два вида с идентичными экологическими потребностями обычно разобщены в пространстве или во времени: они живут в разных биотопах, в разных ярусах леса, обитают в одном водоеме на разных глубинах и т.д. В качестве примера конкурентного исключения можно привести изменение численности плотвы, красноперки и окуня при их совместном проживании в озерах. Плотва со временем вытесняет красноперку и окуня. Исследования показали, что конкуренция сказывается на стадии мальков, когда кормовые спектры молоди перекрываются. В это время мальки плотвы оказываются более конкурентоспособными. В природе конкурирующие за пищу или пространство виды часто избегают или снижают конкуренцию переселением в другое местообитание с приемлемыми для себя условиями, либо переходом на более труднодоступную или трудноусваиваемую пищу, либо сменой времени (места) добычи корма. Происходит деление животных на дневные и ночные (ястребы и совы, ласточки и летучие мыши, кузнечики и сверчки, различные виды рыб, проявляющие активность в разное время суток); львы охотятся на более крупных животных, а леопарды — на более мелких; для тропических лесов характерно распределение животных и птиц по ярусам. Примером разделения жизненного пространства может служить раздел сфер питания между двумя видами баклана — большим и длинноносым. Они обитают в одних и тех же водах и гнездятся на одних и тех же обрывах. Наблюдения показали, что длинноносый баклан ловит рыбу, плавающую в верхних слоях воды, тогда как большой баклан добывает пищу в основном у дна, где ловит камбалу и лонных беспозвоночных. Пространственное разделение можно наблюдать и среди растений. Произрастая совместно в одном местообитании, растения простирают свои корневые системы на разную глубину, тем самым разделяя области поглощения питательных элементов и воды. Глубина проникновения может колебаться от нескольких миллиметров у корнеподстилочных растений (таких как кислица) до десятков метров у крупных деревьев. · 15)Естественный отбор. Формы естественно отбора. Искусственный отбор. Естественный отбор — основной эволюционный процесс, в результате действия которого в популяции увеличивается число особей, обладающих максимальной приспособленностью (наиболее благоприятными признаками), в то время, как количество особей с неблагоприятными признаками уменьшается. В свете современной синтетической теории эволюции естественный отбор рассматривается как главная причина развития адаптаций, видообразования и происхождения надвидовых таксонов. Естественный отбор — единственная известная причина адаптаций, но не единственная причина эволюции. К числу неадаптивных причин относятся генетический дрейф, поток генов и мутации. Термин «Естественный отбор» популяризовал Чарльз Дарвин, сравнивая данный процесс с искусственным отбором, современной формой которого является селекция. Идея сравнения искусственного и естественного отбора состоит в том, что в природе так же происходит отбор наиболее «удачных», «лучших» организмов, но в роли «оценщика» полезности свойств в данном случае выступает не человек, а среда обитания. К тому же, материалом как для естественного, так и для искусственного отбора являются небольшие наследственные изменения, которые накапливаются из поколения в поколение. отбора[править | править вики-текст |
||||||||||||||
|
|
Последнее изменение этой страницы: 2016-06-26; просмотров: 597; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.216.77 (0.015 с.) |


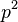 — доля гомозигот по одному из аллелей;
— доля гомозигот по одному из аллелей;  — частота этого аллеля;
— частота этого аллеля;  — доля гомозигот по альтернативному аллелю;
— доля гомозигот по альтернативному аллелю; 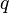 — частота соответствующего аллеля;
— частота соответствующего аллеля;  — доля гетерозигот.Процесс наследования не влияет сам по себе на частоту аллелей в популяции, а возможные изменения её генетической структуры возникают вследствие других причин. Закон действует в идеальных популяциях, состоящих из бесконечного числа особей, полностью панмиктических и на которых не действуют факторы отбора. В медицинской генетике закон Харди — Вайнберга позволяет оценить популяционный риск генетически обусловленных заболеваний, поскольку каждая популяция обладает собственным аллелофондом и, соответственно, разными частотами неблагоприятных аллелей. Зная частоты рождения детей с наследственными заболеваниями, можно рассчитать структуру аллелофонда. В то же время, зная частоты неблагоприятных аллелей, можно предсказать риск рождения больного ребёнка. Основные положения СТЭвыводятся как следствия из закона Харди-Вайнберга. Известно, что пониманиесущности и значения закона вызывает у школьников затруднение, хотя егоматематический аппарат прост и доступен всем, кто знаком с алгеброй среднейшколы. Важно сосредоточить внимание учащихся не только на определении закона -частоты генов и генотипов в популяции не меняются в ряду поколений, - егоусловиях - бесконечно большая популяция, случайное свободное скрещиваниеособей, отсутствие мутационного процесса, естественного отбора и другихфакторов, - математической модели - AA p2 + Aa 2 p + aaq2 = 1, - но и на практическом применении закона.
— доля гетерозигот.Процесс наследования не влияет сам по себе на частоту аллелей в популяции, а возможные изменения её генетической структуры возникают вследствие других причин. Закон действует в идеальных популяциях, состоящих из бесконечного числа особей, полностью панмиктических и на которых не действуют факторы отбора. В медицинской генетике закон Харди — Вайнберга позволяет оценить популяционный риск генетически обусловленных заболеваний, поскольку каждая популяция обладает собственным аллелофондом и, соответственно, разными частотами неблагоприятных аллелей. Зная частоты рождения детей с наследственными заболеваниями, можно рассчитать структуру аллелофонда. В то же время, зная частоты неблагоприятных аллелей, можно предсказать риск рождения больного ребёнка. Основные положения СТЭвыводятся как следствия из закона Харди-Вайнберга. Известно, что пониманиесущности и значения закона вызывает у школьников затруднение, хотя егоматематический аппарат прост и доступен всем, кто знаком с алгеброй среднейшколы. Важно сосредоточить внимание учащихся не только на определении закона -частоты генов и генотипов в популяции не меняются в ряду поколений, - егоусловиях - бесконечно большая популяция, случайное свободное скрещиваниеособей, отсутствие мутационного процесса, естественного отбора и другихфакторов, - математической модели - AA p2 + Aa 2 p + aaq2 = 1, - но и на практическом применении закона.


