
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Порівняльна характеристика чоловічих і жіночих статевих клітинСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
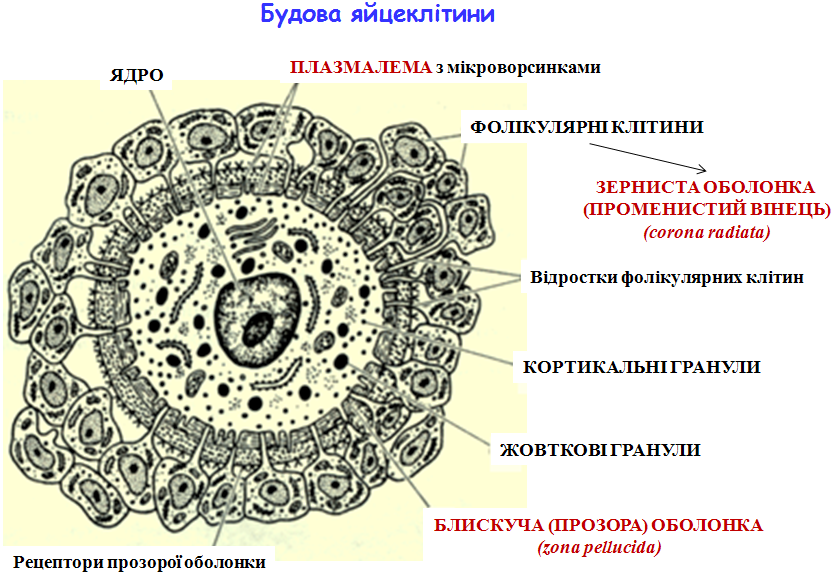
Будова статевих клітин Статеві клітини, або гамети — це клітини з гаплоїдним набором хромосом, які виконують функцію передачі спадкової інформації від особин батьківського покоління нащадкам.На відміну від нестатевих клітин, гамети характеризуються низьким рівнем процесів обміну речовин. Яйцеклітини – жіночі статеві клітини. ü здебільшого нерухливі, мають кулясту або овальну форму; ü мають значно більші розміри, ніж соматичні клітини і сперматозоїди (d яйцеклітини ссавців — 100-200 мкм); ü
Особливості будови яйцеклітини: 1. Оболонки: - зерниста оболонка утворена фолікулярними клітинами, відростки яких проникають у блискучу оболонку, утворюючи „променистий вінець” (corona radiata) (живлення овоцита) - блискуча, або прозора оболонка (zona pellucida) утворена глікопротеїнами zp 1, zp 2, zp 3. Це складна система біологічного захисту овоцита, що забезпечує видоспецифічне зв’язування з найбільш „якісним” сперматозоїдом. 2. Перивітеліновий простір (між плазмалемою та блискучою оболонкою). Тут знаходяться полярні тільця (продукти мейозу, що мають n-набір хромосом, але практично позбавлені цитоплазми). 3. Плазмалема може утворювати мікроворсинки. 4. Специфічні структури цитоплазми: - кортикальні гранули знаходяться під плазмалемою, містять ферменти, що після запліднення беруть участь у кортикальній реакції; - жовткові гранули оточені мембраною та містять фосфопротеїн фосфовітин та ліпопротеїни ліповітелін. 5. Цитоплазма овоцита характеризується високим вмістом компонентів білоксинтезуючої системи. 6. У людини до моменту запліднення ядро яйцеклітини знаходиться на стадії метафази ІІ мейотичного поділу, тому правильно говорити, що запліднюється овоцит ІІ порядку.
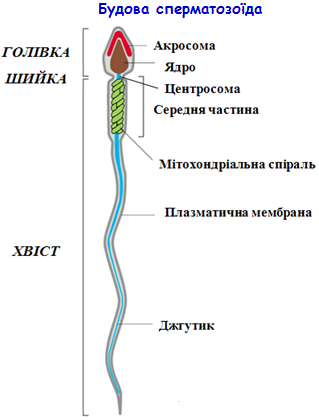
ü здебільшого рухливі клітини, які мають видовжену форму тіла; ü розміри завжди мікроскопічні (у ссавців 50-60 мкм); ü типові сперматозоїди мають головку, шийку і джгутик;
ü у частини вищих рослин, деяких водоростей і грибів сперматозоїди джгутиків не мають; ü усі сперматозоони мають негативний заряд, що перешкоджає їх склеюванню; ü утворюються в спеціалізованих органах: у вищих рослин — в антеридіях, у тварин — у сім'яниках.
Особливості будови сперматозоїда:
1. Голівка: - плазмалема містить білки, що беруть участь у таксисі та зв’язуванні з яйцеклітиною; - ядро (дуже ущільнене, без гістонових білків, з n-набором хромосом); - акросома – обмежений мембраною сплющений пухирець з гідролітичними ферментами. Це видозмінений апарат Гольджі, а функціонально – це велика лізосома. 2. Шийка коротка і містить пару центріолей, що лежать взаємоперпендикулярно.
3. Тіло (проміжний відділ) містить: - аксонему (осьову нитку джгутика), що є результатом видовження мікротрубочок однієї з центріолей і проходить вздовж всієї наступної частини джгутика; - мітохондрії, що у вигляді спіралі оточують джгутик; - 9 жорстких фібрил між аксонемою та мітохондріальною спіраллю.
4. Хвіст містить аксонему, оточену фібрилярним футляром. У кінцевій частині хвоста є лише аксонема, оточена плазмалемою.
Порівняльна характеристика чоловічих і жіночих статевих клітин
Процеси формування статевих клітин Гаметогенез — процес утворення і дозрівання статевих клітин, який відбувається в статевих залозах. Гамети в багатоклітинних тварин розвиваються в статевих залозах – гонадах, у рослин і грибів – у гаметангіях. ЕТАПИ СПЕРМАТОГЕНЕЗУ
ОВОГЕНЕЗ
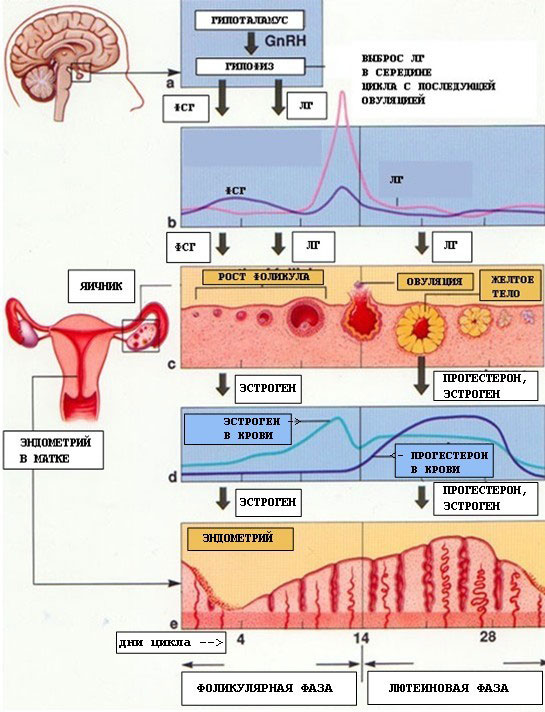
МЕНСТРУАЛЬНИЙ ЦИКЛ
Ще в кінці ХІХ ст. вітч. вчені Д.О. Отт, С.С. Жихарєв, А.В. Репрєв встановили, що менструальний цикл не є місцевим процесом, а хвилеподібною реакцією організму, пов’язаною із змінами в системі гіпоталамус-гіпофіз-яєчники-матка, що ззовні проявляється матковою кровотечею. Ці зміни важливих життєвих процесів в організмі жінки вони назвали "менструальною хвилею". Віковий період жінки, коли проявляється здатність до запліднення і народження дитини, називають репродуктивним. Він тісно пов’язаний з появою фізіологічного ритму в функції жіночого орагнізму, зовнішнім проявом якого є місячні (menses).
Кожен нормальний менструальний цикл є підготовкою організму жінки до вагітності. Запліднення і вагітність настають переважно в середині менструального циклу після овуляції (розриву зрілого фолікула) і виходу готової до запліднення яйцеклітини. Якщо в цей період не відбувається запліднення, то незапліднена яйцеклітина гине, а підготовлена для неї слизова оболонка матки відшаровується, починається менструальна кровотеча. Таким чином, поява менструації свідчить про закінчення складних циклічних змін в організмі жінки, спрямованих на підготовку до можливого настання вагітності, і,разом із тим, про початок нового циклу. В клінічній практиці початком менструального циклу прийнято вважати 1-й день менструації, а тривалість кожного циклу визначають від початку одієї до початку другої менструації.
Нормальна тривалість менструального циклу коливається від 21 до 35 днів і в більшості випадків складає 28 днів. Тривалість самої менструації 2-7 днів. При цьому відокремлюється функціональний шар слизової оболонки матки. Крововтрата в середньому складає приблизно 50-100 мл. Менструація у дівчаток починається в 12-14 років. Першу менструацію називають menarche. Репродуктивна система жінки досягає оптимальної функціональної активності в 17-18 років і безперервно діє протягом 23-28 років репродуктивного періоду жінки. Регуляція менструальної функції проходить складним нейрогуморальним шляхом. Циклічні зміни в організмі жінки, пов’язані із здійсненням менструальної функції, відбуваються при обов’язковій участі п’яти ланок, або рівнів регуляції. Кожна із них регулюється вищерозташованими структурами за механізмом зворотного зв’язку. Рівні (ланки) регуляції менструальної (репродуктивної) функції: 1. Кора головного мозку - церебральні структури. В корі головного мозку ще не визначено точну локалізацію центрів, що регулюють менструальну функцію. Припускають, що імпульси із зовнішнього середовища та інтерорецепторів внутрішнього середовища через систему передавачів - нейротрансміттерів надходять в нейросекреторні ядра гіпоталамуса. Виділено і синтезовано класичні синаптичні нейротрансміттери: біогенні аміни - катехоламіни (дофамін, норадреналін, серотонін, індол) і клас морфіноподібних опіатів. Порушення репродуктивної функції жінки реалізуються через зміни синтезу нейротрансміттерів в нейронах мозку з подальшими змінами в гіпоталамусі. 2. Гіпоталамус - підкоркові центри. Гіпоталамус є центральним регулятором менструальної функції. Він є передавальним пунктом між імпульсами, що надходять через ЦНС із зовнішнього середовища, і впливом гормонів залоз внутрішньої секреції та внутрішнього середовища. Ядра нервових клітин гіпоталамуса виділяють нейросекрети – релізинг-гормони, що належать до низькомолекулярних поліпептидів. Нейрогормони гіпоталамусу, які стимулюють продукцію тропних гормонів гіпофізу, називають ліберинами, а ті, що її гальмують – статинами. Гіпоталамус виробляє 3 релізинг-факторів, які мають пряме відношення до регуляції менструальної функції і сприяють вивільненню в передній долі гіпофізу відповідних тропних гормонів: фолікулостимулюючий релізинг-фактор (ФСГ-РФ), або фоліберин; лютеїнізуючий релізинг-фактор (ЛРФ), або люліберин; пролактинвивільняючий релізінг-фактор (ПРФ), або пролактоліберин. З їх допомогою відбувається вивільнення в аденогіпофізі трьох відповідних гормонів, які називають гонадотропінами.
З факторів, які пригнічують вивільнення в аденогіпофізі тропних гормонів (статинів), безпосереднє відношення до регуляції менструальної функції має пролактинінгібуючий фактор (ПІФ), або пролактостатин. Секреція нейрогормонів здійснюється як безперервно на відносно низькому рівні, так і циклічно відповідно до фаз менструального циклу. 3. Гіпофіз – ендокринна залоза. Система гіпоталамус - гіпофіз являє собою єдиний функціональний комплекс, що відіграє важливу роль в регуляції репродуктивної і менструальної функції. Релізинг-гормони із гіпоталамуса по аксонах нервових клітин потрапляють в судини портальної кровоносної системи, яка тісно з’єднана з гіпофізом. Особливістю цієї системи є можливість току крові в обидві сторони як до гіпоталамусу, так і до гіпофізу, що досить важливо для механізму зворотного зв’язку між цими структурами. Регуляцію репродуктивної (менструальної) функції жінки здійснюють три гонадотропних гормони аденогіпофізу: - фолітропін, або фолікулостимулюючий гормон (ФСГ) – стимулює ріст фолікулів; - лютропін, або лютеінізуючий гормон (ЛГ) – стимулює синтез прогестерону в жовтому тілі яєчника після овуляції; - пролактин – стимулює ріст молочних залоз, сприяє регуляції лактації, здійснює жиромобілізуючий ефект. Збільшення секреції пролактину призводить до порушення стероїдогенезу в яєчниках, ановуляції і неплідності. Яєчники - статеві залози. Овуляція настає при синергічній дії ФСГ та ЛГ, при домінуючій дії останнього. Протягом всього менструального циклу гіпофізом виділяються обидва гонадотропіни, тільки в різній кількості. З початком менструального циклу збільшується секреція ФСГ, що призводить до стимуляції і росту фолікулів в яєчниках. ФСГ і ЛГ доводять розвиток фолікула до стадії зрілості. За кілька годин до овуляції настає різкий максимум – овуляторний пік виділення ЛГ, що дає поштовх до овуляції. В 2 фазу циклу виділяється в більшій кількості ЛГ, який сприяє продукції прогестерону жовтим тілом в яєчнику. Циклічність функції гіпоталамусу, гіпофізу і яєчників складає гіпоталамо-гіпофізарно-яєчниковий комплекс, наслідком дії якого є репродукція і менструація жінки. Яєчниковий (оваріальний) цикл має 2 фази: перша (фолікулінова) - з першого дня циклу і до моменту овуляції, друга (лютеїнова) - від овуляції до першого дня наступної менструації. В першу фазу циклу під впливом ФСГ і ЛГ гіпофізу починає рости і дозрівати фолікул. За життя жінки основна маса фолікулів атрофується і тільки 10 % досягають зрілості, проходять цикл розвитку від примордіального до преовуляторного фолікула, овулюють і перетворюються в жовте тіло. З настанням статевої зрілості під впливом гонадотропних гормонів аденогіпофізу навколо овоцитів розростаються так звані зернисті клітини і утворюється вторинний фолікул (малий зрілий фолікул). Подальший розвиток вторинного фолікула призводить до утворення третинного, або везикулярного, фолікула, який містить порожнину, заповнену рідиною, і овоцит, що являє собою найбільшу клітину в організмі жінки діаметром 0,1 мм. Внутрішня поверхня фолікула покрита кількома рядами гранульозних клітин, які під впливом гонадотропінів гіпофізу продукують стероїдні гормони.
Періодично, відповідно до тривалості менструального циклу, відбувається дозрівання чергового фолікула. Всього за весь репродуктивний період життя жінки дозріває приблизно 400 фолікулів. Примордіальний фолікул складається з яйцеклітини, яка оточена рядом сплощених клітин фолікулярного епітелію. При дозріванні фолікула яйцеклітина збільшується в розмірах, клітини фолікулярного епітелію розмножуються і округлюються, утворюючи зернистий шар фолікула. Вони секретують рідину, яка містить естрогени. Ця рідина відтісняє яйцеклітину до периферії. Яйцеклітина, відтіснена до периферії, оточується 17-50 рядами гранульозних клітин. Виникає яйценозний горбик (cumulus oophorus). Фолікул тепер називають граафовим пухирцем. Тиск в середині фолікула підвищується, і при комбінованій дії гонадотропінів та рефлекторній дії окситоцину і протеолітичних ферментів його стінка в ділянці вільного краю розривається – настає овуляція. Незадовго до овуляції відбувається перший мейоз. Після овуляції яйцеклітина з черевної порожнини потрапляє в маткову трубу, в ампулярній частині якої проходить другий редукційний поділ (другий мейоз). Після овуляції під впливом переважаючої дії ЛГ спостерігаються подальше розростання гранульозних клітин і сполучнотканинних оболонок фолікула і накопичення в них ліпідів, що призводить до утворення жовтого тіла. Починається друга лютеїнова фаза. Процес розвитку жовтого тіла ділять на чотири фази: проліферації, васкуляризації, розквіту і зворотного розвитку. При настанні вагітності жовте тіло продовжує розвиватися до 16 тижнів вагітності. В гранульозних клітинах фолікула під впливом ФСГ і ЛГ виробляються стероїдні статеві гормони (естрогени, гестагени і андрогени). Естрогени (грец. оеstros – жага, бажання) головним чином, виділяються клітинами гранульозного (зернистого) шару, в меншій кількості в жовтому тілі фолікула і в корі надниркових залоз. Під впливом естрогенів у дівчаток розвиваються вторинні статеві ознаки у вигляді типового для жінок розподілу підшкірножирового шару, характерної форми тазу, збільшення молочних залоз, росту волосся на лобку і в підпахвинних ділянках. Крім того, естрогени сприяють росту і розвитку статевих органів, особливо матки, під їх дією ростуть малі статеві губи, подовжується піхва, змінюється характер секреції каналу шийки матки, що дає можливість сперматозоїдам легко проникати через слизову пробку в матку. Естрогени викликають проліферацію ендометрію, стимулюють регенерацію ендометрію після менструації, прискорюють гіпертрофію і гіперплазію міометрію під час вагітності, покращують матково-плацентарний кровообіг, сенсибілізують міометрій до середників, що скорочують матку. Під впливом естрогенів в ампулярній частині маткових труб поліпшуються кровопостачання, м’язова активність, посилюється перистальтика маткових труб в період овуляції і міграції заплідненої яйцеклітини. Естрогени чинять значний вплив на терморегуляцію і обмінні процеси. Гестагени (грец. gesto – носити, бути вагітною) сприяють нормальному розвитку вагітності, відіграють велику роль в циклічних змінах ендометрію. Місцем утворення гестагенів в організмі є жовте тіло, клітини зернистого шару фолікулів в яєчниках. В невеликій кількості гестагени синтезуються в корі надниркових залоз. Прогестерон як основний гестагенний гормон також спричиняє виражену біологічну дію на органи мішені: статеві органи, молочні залози і гіпофіз. Під впливом гестагенів пригнічуються збудливість і скоротлива здатність міометрію, збільшуються його розтяжимість і пластичність. Гестагени сприяють підготовці молочних залоз до лактації, діючи на альвеолярний апарат молочних ходів, засвоєнню організмом речовин, зокрема білків, поряд з анаболічним ефектом зумовлюють незначне підвищення температури тіла, особливо базальної. Андрогени (грец. andros – чоловік) – це статеві чоловічі гормони, що в надлишковій кількості викликають у жінки ознаки маскулінізації. Основне джерело утворення андрогенів у жінки - кора надниркових залоз. Невелика їх кількість виділяється інтерстиціальними клітинами і внутрішньою оболонкою фолікулів. Андрогени в організмі затримують азот, натрій, хлор, прискорюють ріст кісток. Вони виробляються в невеликій кількості яєчниками у так званих гілюсних клітинах. Андрогени сприяють росту волосся в аксилярних впадинах і на лобку, а також розвитку клітора і великих статевих губ. Як і гестагени, вони мають анаболічні властивості. Матка - орган – мішень.
Зміни в будові і функції матки в цілому, особливо в будові і функції ендометрію, що настають під дією яєчникових статевих гормонів, називають матковим циклом. Протягом маткового циклу спостерігається послідовна зміна чотирьох фаз циклічних змін в ендометрії: 1) проліферації; 2) секреції; 3) десквамації; 4) регенерації. Фази проліферації і секреції є основними в матковому циклі, межа між ними - овуляція. 1 фаза проліферації ендометрію починається після завершення регенерації слизової оболонки матки, що відшарувалась під час попередньої менструації. Триває з 5 по 14 день циклу. Початок фази пов’язаний із зростаючою дією на слизову оболонку матки естрогенів, які продукуються фолікулом, що зріє. На початку фази проліферації залози ендометрію вузькі і рівні. У міру наростання проліферації залози збільшуються в розмірах і починають злегка звиватися. Максимально виражена проліферація ендометрію проходить до моменту повного дозрівання фолікула і овуляції (до 12-14 дня 28-денного циклу). Товщина слизової оболонки матки досягає 3-4 мм. Ці зміни відбуваються під впливом гормонів яєчників естрогенів. 2 фаза секреції починається під впливом швидко наростаючої активності жовтого тіла і гестагенів, які виробляються яєчниками. Залози ендометрію набувають звивистого характеру і заповнюються секретом, який вони виробляють. Секрет містить глікоген, глікопротеїди, кислі глікозаміноглікани, які є сприятливим середовищем для імплантації плідного яйця. Слизова оболонка розпушується, крізь неї проростають спірально звиті артеріоли. До кінця фази секреції просвіт залоз ендометрію набуває пилоподібної форми, в них виявляється ще більша кількість секрету. В стромі ендометрію в цей час з’являються так звані псевдодецидуальні клітини. Слизова оболонка стає максимально готовою до прийняття заплідненої яйцеклітини на 21-й день циклу, коли жовте тіло в яєчнику досягає свого розквіту. Товщина ендометрію досягає 8-10 мм. На цей час ендометрій чітко розділяється на два шари: поверхневий і глибокий. Після 21-22 дня циклу, в зв’язку з регресом жовтого тіла, яке перетворюється поступово на біле тіло, знижується концентрація гормону жовтого тіла - прогестерону і порушується трофіка ендометрію, наростають дегенеративні зміни: зменшується на 25-30 % висота ендометрію, посилюється складчастість стінок залоз, вони набувають зірчастого характеру, знижується кількість секрету залоз, виявляються вогнищеві крововиливи в строму. Функціональний шар ендометрію готується до розпаду і відшарування. Якщо взяти гістологічний зріз ендометрію в цей час, то можна побачити ці зміни під мікроскопом, їх називають анатомічною менструацією, яка виявляється за добу до явної фізіологічної, або клінічної менструації. 3 фаза – десквамації, або менструація, яка триває в середньому 3-4 дні. Внаслідок різкого зниження рівня всіх статевих гормонів (прогестерону і естрогенів) відбувається відшарування функціонального шару слизової оболонки матки, що клінічно проявляється кровотечею. Для настання менструації в організмі жінки діє ряд факторів: 1. Спад всіх статевих гормонів більше ніж на 50 % за кілька днів до менструації. 2. Судинний спазм атреріол в ендометрії, підвищена проникність стінок судин. 3. Порушення кровообігу, застій і деструктивні зміни в ендометрії. 4. Утворення ділянок некрозу і гематом слизової оболонки ендометрію. 5. Підвищення активності фібринолітичних ферментів та ін. Після повного відторгнення ендометрію на 3-4 день циклу настає четверта фаза регенерації, що здійснюється з строми базального шару слизової оболонки і донних залоз. Вона триває 2-3 дні. Встановлено, що в середині менструального циклу, а точніше до часу настання овуляції, спостерігається чітко виражене розширення зовнішнього вічка шийки матки. Одночасно виявляються пов’язані з овуляцією зміни деяких фізичних та хімічних властивостей слизу, що продукується залозами слизової оболонки шийкового каналу. Так, в міру наростання проліферації кількість слизу в шийковому каналі поступово зростає, він стає більш проникним для світла і розтяжимим (розтягається у вигляді нитки до 8-10 см). Зовнішній вигляд вічка шийки матки нагадує зіницю. Поява симптому "зіниці" свідчить про високу продукцію естрогенів яєчниками. При наближенні терміну овуляції і під впливом зростаючої активності естрогенів одночасно з вказаними вище фізичними властивостями змінюється і хімічна характеристика слизу шийкового каналу. Це проявляється появою так званого симптому арборизації. Після висушування на повітрі шийкового слизу, нанесеного у вигляді мазка на предметне скло, в препараті випадають кристали хлориду натрію. На початку першої і наприкінці другої фаз циклу ці кристали розташовуються аморфно. В більш пізній період І фази і ранній період ІІ фази, особливо до початку овуляції, вони розміщуються у вигляді листка папороті. Описані зміни шийки матки і властивостей шийкового слизу мають велике біологічне значення, оскільки полегшують потрапляння сперматозоїдів через шийковий канал в порожнину матки, а звідти – в маткові труби. Після настання овуляції і утворення жовтого тіла всі перераховані зміни шийки матки і шийкового слизу зазнають зворотнього розвитку: зовнішнє вічко змикається, слиз стає густим, мутним, розтяжимість його знижується, зникає феномен арборизації. ОНТОГЕНЕЗ Онтогенез — це індивідуальний розвиток особини від її зародження до смерті. Тривалість онтогенезу у різних організмів є різною (наприклад, у секвойї понад 3 тис. років) і не залежить від рівня організації. Поява змін в онтогенезі визначається генотипом. Індивідуальний розвиток включає ріст (кількісні зміни) та диференціацію клітин (якісні зміни) і здійснюється під впливом багатьох чинників (наприклад, умови середовища, нервова, гуморальна регуляція та ін.). У різних груп організмів онтогенез має свої особливості, які залежать від рівня організації (в одноклітинних організмів збігається з клітинним циклом), способів розмноження (при вегетативному розмноженні онтогенез зводиться до диференціації клітин і органів багатоклітинного зачатка; при нестатевому розмноженні зародження починається із спори; при статевому розмноженні вихідною стадією є зигота). Запліднення
Завершення мейозу - одночасно з процесами блокування поліспермії завершується ІІ поділ мейозу в овоциті; - це збільшує кількість полярних тілець у перивітеліновому просторі;
Дроблення
– це ряд послідовних мітотичних поділів зиготи, в результаті яких запліднена яйцеклітина – зигота – перетворюється в багатоклітинний комплекс – бластулу. Кількість клітин збільшується внаслідок мітозу, але інтерфаза дуже коротка і бластомери не ростуть (в інтерфазі лише S-період). Особливості дробіння в різних груп організмів залежать від характеру розташування та кількості жовтка, тому виділяють два типи дробіння.
Класифікація яйцеклітин 1.За кількістю жовтка - полі- (багатожовткові; риби, плазуни, птахи); - мезо- (середньожовткові; осетрові, амфібії); - оліго- (маложовткові; більшість червів, молюсків, голкошкірі); - алецитальні (безжовткові яйця). 2.За розміщення жовтка по об’єму яйця: - телолецитальні – кількість жовтка наростає від одного полюса до іншого (риби, плазуни, птахи, осетрові, амфібії); - гомолецитальні (ізолецитальні) – жовткові гранули рівномірно розподілені в яйці (більшість червів, молюски, голкошкірі); - центролецитальні – вільна від жовткових гранул цитоплазма розміщена зразу ж під оболонкою яйця, навколо ядра, яке займає центральне положення, і у вигляді тонких тяжів сполучає названі області. Проміжний простір заповнений жовтком (членистоногі). Класифікація бластул Целобластула – складається з більш або менш однакових бластомерів і великим бластоцелем всередині, утвореним в результаті повного рівномірного дроблення. Амфібластула складається з неоднакових мікромерів і макромерів. Бластоцель невеликий і зсунутий до анімального полюсу. Перибластула – не має бластоціля і утворюється в результаті поверхневого дроблення. Дискобластула являє собою диск бластомерів, які лежать на жовтку, що не роздробився. Бластула у вигляді двохшарової пластинки, що має щілинну порожнину називається плакулою. Морулою іноді називають саму ранню бластулу, коли зародок містить вже досить значну кількість клітин (32-64), але порожнина дроблення ще не сформована.
Утворення бластоцисти Через 4,5 - 5 діб утворюється бластоциста – зародок з порожниною, заповненою рідиною. У вигляді вільної бластоцисти зародок знаходиться в порожнині матки близько 2-х діб – на 5-7 добу. Поділ клітин в бластоцисті зберігає характер асинхронного і нерівномірного.
Компоненти бластоцисти: - трофобласт – одношарова стінка из дрібних світлих клітин (згодом з нього розвивається позазародковий орган – хоріон); - ембріобласт (внутрішня клітинна маса) – сукупчення великих темних бластомерів у вигляді вузлика на внутрішній поверхні трофобласта біля одного з полюсів; - бластоцель – порожнина, заповнена рідиною.
Ріст бластоцисти - завдяки всмоктуванню трофобластом рідини із порожнини матки, об’єм пухирця дещо збільшується; у бластомерах все більше активуються синтетичні процеси; - у трофобласті з’являються вирости, які поступово руйнують оболонку запліднення навколо зародка; у результаті, за кілька годин до імплантації зародок втрачає цю оболонку («вилуплювання», або хетчинг бластоцисти);
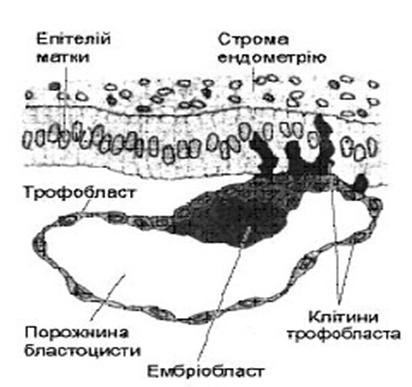
Імплантація – це занурення зародка в товщу ендометрія (слизової оболонки матки). Вона починається на 6-7 добу ембріогенезу і триває 40 годин. Звичайне місце імплантації – верхня частина матки, передня або задня стінка.
Гаструляція — це процес формування двошарового зародка — гаструли. Ріст клітин при гаструляції не відбувається. На цьому етапі утворюються два або три шари тіла зародка — зародкові листки. У процесі гаструляції необхідно розрізняти два етапи: - утворення екто- та ентодерми (формується рання гаструла — двошаровий зародок);
На етапі гаструляції завершується ембріогенез двошарових тварин (губки, кишковопорожнинні), мезодерма закладається в зародковому розвитку тришарових тварин (починаючи з плоских червів). Способи гаструляції, тобто механізми утворення зародкових листків, відрізняються у різних тварин і визначаються будовою бластули. Існує чотири основні способи гаструляції: - інвагінація (вп’ячування) – в середину бластоцеля входять не окремі клітини, а ціла ділянка клітинного пласту, яка не втрачає епітеліальної структури (сцифоїдні медузи, коралові поліпи); - делямінація (розшарування) – зустрічається у кишковопорож-нинних, де дроблення завершується морулою без внутрішньої порожнини. Переміщення клітин практично відсутні; - іміграція (виселення) – відкритий І.І.Мечніковим у 1886р. – це вселення у порожнину бластоцелю окремих клітин з стінки бластули (у деяких гідромедуз). Найбільш еволюційно древній; - епіболія (обростання) – інвагінація іноді неможлива через малі розміри бластоцелю та інертність великих бластомерів (у малощетинкових червів тільки епіболія). З процесу гаструляції починаються ембріональна диференціація і морфогенез. Період гаструляції характеризується переміщенням окремих клітин, груп клітин і клітинних пластів, погодженими змінами клітинної форми, діленням клітин, контактними взаємодіями між клітинами, виселенням і вселенням клітин, що призводить до розчленування зародка на зародкові листки: ектодерму, мезодерму, ентодерму. Зародкові листки у різних тварин є гомологічними утворами, тобто в ході розвитку вони дають ідентичні структури. В результаті гаструляції виникає зародок – гаструла. Гаструла має порожнину – гастроціль (порожнина первинної кишки), в яку веде отвір – бластопор (первинний рот). В залежності від подальшої долі бластопора в розвитку всі тварини підрозділяються на первиннороті і вториннороті. У первинноротих, до яких відноситься більшість безхребетних, на місці бластопора утворюється ротовий отвір, а протилежний кінець стає заднім кінцем тіла. У вторинноротих, які включають хордових і деяких безхребетних, бластопор перетворюється в анальний отвір, або в особливий нервовокишечний канал, розміщений на задньому кінці тіла зародка, а ротовий отвір проривається на черевному боці на протилежному кінці тіла.
ГІСТОГЕНЕЗ ТА ОРГАНОГЕНЕЗ Гістогенез – розвиток та утворення тканин. В основі гістогенезу лежить процес диференціювання клітин, що приводить до їх спеціалізації, яка виражається в появі у клітин специфічних ознак і виконанню клітинами певних функцій. Одночасно з гістогенезом відбувається і органогенез – розвиток органів. При цьому темпи гісто- і органогенезу можуть не співпадати. У багатьох органах тканинне диференціювання закінчується тільки після народження. Орган може набувати певної форми вже після того, як закінчилась диференціація тканин. Здійснюються ці процеси внаслідок диференціації (виникнення відмінностей у будові та функціях клітин, тканин, органів). У гістогенезі рослин беруть участь ініціальні клітини твірних тканин, а в гістогенезі тварин — стовбурні, напівстовбурні та дозрілі клітини. Велике значення при органогенезі відіграють міжклітинні взаємодії, вплив біологічно активних речовин.
Фазами гістогенезу та органогенезу (на прикладі ланцетника) є нейруляція — утворення осьового комплексу органів (нервової трубки, хорди), формування інших орган |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 18212; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.116.89.70 (0.016 с.) |

 Сперматозоїди – чоловічі статеві клітини.
Сперматозоїди – чоловічі статеві клітини.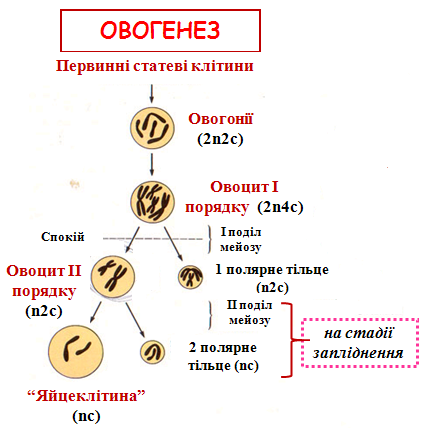


 - саме у цей короткий період овоцит ІІ є яйцеклітиною
- саме у цей короткий період овоцит ІІ є яйцеклітиною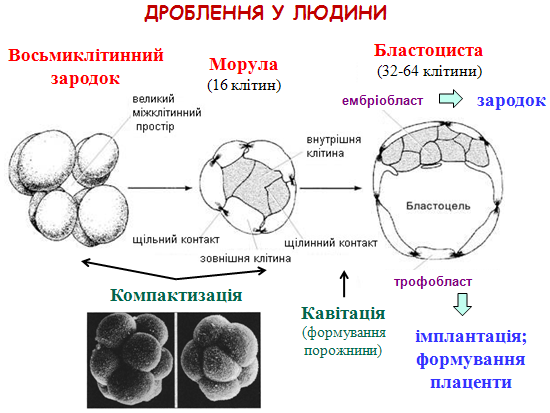
 - після цього оболонка вже не заважає зародку збільшуватися у розмірах, клітинні цикли стають звичайними, з повноцінною інтерфазою і ростом клітин; тому в подальшому збільшення маси зародка відбувається значно швидше; окрім цього хетчинг дозволяє «оголити» рецептори бластомерів, що робить можливими процеси взаємодії між клітинами зародка і клітинами ендометрію матки, що лежать в основі імплантації.
- після цього оболонка вже не заважає зародку збільшуватися у розмірах, клітинні цикли стають звичайними, з повноцінною інтерфазою і ростом клітин; тому в подальшому збільшення маси зародка відбувається значно швидше; окрім цього хетчинг дозволяє «оголити» рецептори бластомерів, що робить можливими процеси взаємодії між клітинами зародка і клітинами ендометрію матки, що лежать в основі імплантації.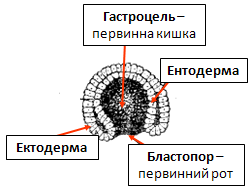 В імплантації розрізняють 2 стадії: адгезії (прилипання) та інвазії (проникнення). На стадії адгезии зародок за допомогою трофобласта прикріплюється до ендометрія. Основною за тривалістю є стадія інвазії. Після адгезії клітини трофобласта починають швидко ділитися і виділяти ферменти, які руйнують прилеглі ділянки ендометрію – епітелій і сполучну тканину. Завдяки цьому зародок поступово занурюється в ендометрій, утворюючи імплантаційну ямку, і, зрештою, повністю опиняється в товщі ендометрію. Одночасно із зануренням зародка, його трофобласт розподіляється на 2 шари: внутрішній – цитотрофобласт, що зберігає клітинну будову і зовіншній – симпластотрофобласт – багатоядерний утвір, що утворений у результаті злиття великої кількості клітин. Впродовж кількох днів зародок живиться через трофобласт продуктами розпаду ендометрію (гістіотрофний тип живлення). Після повного занурення зародка дефект в ендометрії заростає загоюється ндо 12-13 дня. Навколо зародка утворюються лакуни, заповнені материнською кров’ю.
В імплантації розрізняють 2 стадії: адгезії (прилипання) та інвазії (проникнення). На стадії адгезии зародок за допомогою трофобласта прикріплюється до ендометрія. Основною за тривалістю є стадія інвазії. Після адгезії клітини трофобласта починають швидко ділитися і виділяти ферменти, які руйнують прилеглі ділянки ендометрію – епітелій і сполучну тканину. Завдяки цьому зародок поступово занурюється в ендометрій, утворюючи імплантаційну ямку, і, зрештою, повністю опиняється в товщі ендометрію. Одночасно із зануренням зародка, його трофобласт розподіляється на 2 шари: внутрішній – цитотрофобласт, що зберігає клітинну будову і зовіншній – симпластотрофобласт – багатоядерний утвір, що утворений у результаті злиття великої кількості клітин. Впродовж кількох днів зародок живиться через трофобласт продуктами розпаду ендометрію (гістіотрофний тип живлення). Після повного занурення зародка дефект в ендометрії заростає загоюється ндо 12-13 дня. Навколо зародка утворюються лакуни, заповнені материнською кров’ю.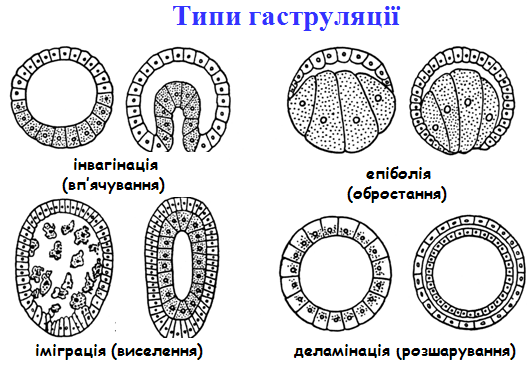 - утворення мезодерми (формується пізня гаструла — тришаровий зародок).
- утворення мезодерми (формується пізня гаструла — тришаровий зародок).


