Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Стадії реплікації: ініціація, елонгація, термінація (на прикладі E.coli)Содержание книги
Поиск на нашем сайте
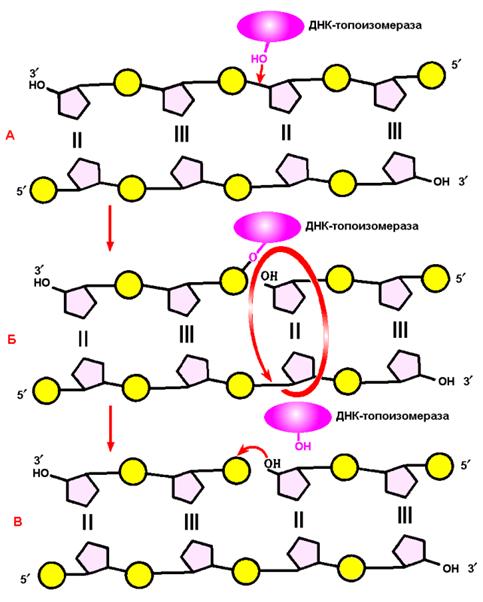
Для передачі дочірнім клітинам генетичної інформації у процесі реплікації ДНК (DNA) має бути створена копія геному. У більшості організмів реплікація здійснюється як двонаправлений процес. У деяких бактеріальних плазмід і вірусів може відбуватися однонаправлена реплікація за типом розгортання рулону або кільця, що котиться. У реплікації так само, як і в процесах транскрипції і трансляції виділяють три основні стадії: ініціацію, елонгацію та термінацію. Розглянемо процес реплікації на прикладі Е. coli. Ініціація починається з перевірки ДНК на цілісність шляхом негативного суперскручування за допомогою ДНК-гірази. Якщо в ДНК є хоч один розрив, суперскручування не відбувається, необхідна репарація ДНК. Для переводу ДНК в релаксований стан (це важливо для розкручування), гіраза робить дволанцюгові розриви в ДНК і розплутує петлі. Далі білок-ініціатор (dnaA) прикріплює збагачену АТ-парами область оріджина до виросту цитоплазматичної мембрани. Топоізомераза І робить одноланцюговий розрив поблизу ori C, прикріплюється до 5’-кінця розриву (рис.8), а 3’-кінець починає розкручуватися відносно інтактного ланцюга з утворенням реплікативної вилки, де й відбувається складання ферментативного апарату реплікації (рис.9). Хеліказа, розділяючи спірально закручені ланцюги материнської ДНК, викликає появу супервитків (тобто сильну спіралізацію молекули ДНК) перед репликаційною виделкою. Супервитки ДНК виникають тому, що при розходженні кожних 10 пар нуклеотидів, які утворюють один виток спіралі, материнська ДНК повинна зробити один повний оберт навколо своєї вісі. Отже, для просування репликаційної вилки вперед вся молекула ДНК перед нею мала б швидко обертатися. Таке обертання вимагало б великих витрат енергії. Насправді ж обертання не відбувається завдяки дії особливого ферменту – ДНК-топоізомерази. Топоізомераза розриває один з ланцюгів ДНК і здійснює обертання навколо іншого (цілого) ланцюга, як навколо вісі обертання (рис. 8). Це усуває супервитки і послаблює напругу, що нагромадилася в подвійній спіралі ДНК. Після зняття надмірної напруги фермент відновлює розірваний ланцюг.
Рис. 8. - Розрив одного з ланцюгів ДНК за допомогою фермента ДНК-топоізомерази Примітка: А–ДНК-топоізомераза утворює ковалентний зв'язок з однією з фосфатних груп ДНК (верхній ланцюг); Б – у результаті розриву фосфодиефірного зв'язку в одному полінуклеотидному ланцюзі навколо відповідного йому зв'язку іншого ланцюга здійснюється обертання, яке знімає напруженість, спричинену розходженням двох ланцюгів ДНК в області реплікаційної вилки; В– після зняття напруги в спіралі ДНК відбувається спонтанне відділення ДНК-топоізомерази та відновлення фосфодиефірного зв'язку в ланцюзі ДНК. Реплікація у прокаріотів починається з ділянки нуклеїнової кислоти, яка називається точкою ініціації реплікації або оріджином (локус ori, від англ.: origin), де два ланцюги батьківської ДНК відокремлюються один від одного; в місцях розходження полінуклеотидних ланцюжків починається синтез дочірніх ланцюгів. Область спіралі ДНК, яка на початку реплікації розкручується в першу чергу, називають ділянкою початку реплікації. На певній відстані від точки ініціації молекула ДНК містить ділянку, у якому реплікація зупиняється. Ця ділянка ДНК носить назву точки термінації (тобто точки закінчення) реплікації. Ділянку ДНК від точки ініціації реплікації до точки її закінчення утворює одиницю реплікації – реплікон.
На лідируючому ланцюзі неспецифічна РНК-полімераза здійснює синтез невеликої РНК-місткого транскрипту, до якого в подальшому ДНК-полимераза ІІІ буде приєднувати ДНК-місткі нуклеотиди (якщо транскрипт утворився в «неправильному» місці, РНК-аза Н гідролізує його, забезпечуючи вибірковість синтезу). Перед транскриптом на ланцюг ДНК «сідає» хеліказа гер та ЅЅВ-білки. Останні утримують розплетені ланцюги ДНК, не даючи їм ренатурувати, а також прибирають випадкові елементи матричного ланцюга ДНК. Далі приєднується до ланцюга ДНК-полімераза ІІІ. На відстаючому ланцюзі формується складна хеліказа-dnaBdnaC шляхом послідовного приєднання шести білків dnaC та шести білків-dnaB. Сюди ж долучаються білки та праймаза. Формується складна частка – праймосома, яка в комплексі з ДНК-полімеразою ІІІ утворює реплісому. На цей ланцюг також «сідають» ЅЅВ-білки. Перед початком елонгації на кожному ланцюзі виявляється по одній молекулі ДНК-полімерази ІІІ. Ще на початку 70-х років ХХ століття Б. Альбертс і А. Корнберг висловили припущення, що редуплікацію ДНК здійснює комплекс білків - реплісома, аналогічний рибосомі. Крім ДНК-полімераз і вже перелічених факторів до складу реплісоми, що формується в реплікаційній вилці, входять також інші білки, які прямо чи побічно приймають участь у синтезі ДНК. Елонгація здійснюється по-різному на лідируючому та відстаючому ланцюгах. Внаслідок відмінностей в механізмах синтезу копій на двох різних ланцюгах материнської ДНК репликативна вилка асиметрична. З двох синтезованих дочірніх ланцюгів один будується безперервно, його синтез йде швидше, тому цей ланцюг називають провідним (лідируючим). Синтез іншого ланцюга йде повільніше, тому що він збирається з окремих фрагментів, які потребують багаторазового формування, а потім видалення РНК-затравки. Тому цей ланцюг називають запізнілим (відстаючим). Хоча на ньому окремі фрагменти утворюються в напрямку 5'→3', в цілому цей ланцюг росте в напрямку 3'→5'. На лідируючому ланцюзі йде безперервний синтез ланцюга ДНК, шляхом послідовного приєднання нуклеотидів до РНК-транскрипту. Звільнені водневі зв'язки нуклеотидів двох роз'єднаних батьківських ланцюгів служать своєрідними магнітами, що притягають з нуклеоплазми вільні нуклеотиди, які знаходяться в нуклеоплазмі у вигляді дезоксирибонуклеозидтрифосфатів: дАТФ, дГТФ, дЦТФ, дТТФ. Комплементарний нуклеозидтрифосфат утворює водневі зв'язки з комплементарною основою ланцюга материнської ДНК. Потім за участю ферменту ДНК-полімерази він зв'язується фосфодиефірним зв'язком з попереднім нуклеотидом знову синтезованого ланцюга, віддаючи при цьому неорганічний пірофосфат (від 3’-фосфонуклеозиду відщеплюється пірофосфат), а залишений 5’-фосфорнокислий кінець приєднується до 3’-гидроксилу попереднього нуклеотиду. Синтез здійснюється в напрямку 5’-3’ (рис. 10). ДНК-полімераза приєднує черговий нуклеотид до ОН-групи тільки в 3'-положенні попереднього нуклеотида. Тому ланцюг поступово подовжується на її 3'-кінці. Особливістю ДНК-полімерази є її нездатність почати синтез нового полінуклеотидного ланцюга шляхом простого зв'язування двох нуклеозидтрифосфатів, їй необхідний 3'-ОН-кінець будь-якого полінуклеотидного ланцюга, спареного з матричним ланцюгом ДНК, до якого ДНК-полімераза може лише додавати нові нуклеотиди. Такий полінуклеотидний ланцюг називають праймером або затравкою. Праймером для початку синтезу ланцюга ДНК є коротка послідовність РНК, що утворюється на ДНК за участю ферменту РНК-праймази. Довжина праймерів варіює від 10 до 100 нуклеотидів [Дубінін Н.П.; 1986; Яригін Ст. Н. та ін, 1997 ]. Оскільки ДНК-полімераза здатна здійснювати збірку полінуклеотида тільки в одному напрямку – від 5'- до 3'-кінця, то процес реплікації триває на антипаралельних ланцюгах-матрицях ДНК по-різному. На одній із матриць (3'→5') збірка нової ланцюга відбувається безперервно від 5'- до 3'-кінця і вона поступово подовжується на 3'-кінці. Інша ланцюг, що синтезується на матриці (5'→3'), повинен зростати від 3'- 5'-кінця, але це суперечить напрямку дії ферменту ДНК-полімерази. Встановлено, що синтез другого ланцюга ДНК здійснюється також в напрямку від 5'- до 3'-кінця, але не безперервно, а численними короткими фрагментами, що одержали назву фрагментів Оказакі. Так вони названі на честь японського молекулярного генетика Рейджи Оказакі. На відстаючому ланцюзі праймази спочатку синтезують РНК-місткі праймери – затравки, до яких ДНК-полімераза ІІІ приєднує послідовно ДНК-місткі нуклеотиди з утворенням фрагментів Оказакі (по 1000-2000 нуклеотидів кожен). У результаті дочірніій ланцюг ДНК стає фрагментарним і складається з РНК-містких затравок і ДНК-містких фрагментів Оказакі, що чергуються. Для перекладу всього ланцюга на мову ДНК до процесу підключається ДНК-полімераза І, яка за допомогою своєї 3’-екзонуклеазної активності прибирає РНК-місткі праймери, а за допомогою полімеризуючої активності забудовує утворені прогалини ДНК-місткими нуклеотидами. Всі ДНК-місткі фрагменти зшиваються лігазою. Розпочавшись в точці ініціації, реплікація продовжується доти, поки весь реплікон не буде редуплікований. Кільцеві молекули ДНК прокаріотичних клітин мають один локус ori і являють собою цілком окремі реплікони. Еукаріотичні хромосоми містять велику кількість репліконів. Тому подвоєння молекули ДНК починається в декількох точках, що прискорює процес реплікації великих молекул ДНК еукаріотів. У результаті азотисті основи роз'єднаних ланцюгів стають доступними для зв'язування комплементарними нуклеотидами, які надходять з нуклеоплазми.
Рис. 10.- Приєднання чергового нуклеотида до дочірнього ланцюга ДНК, що синтезується за участю ДНК-полімерази: Ф-Ф – пірофосфат Нині встановлено, що процеси, що йдуть в реплікативній вилці, ще більш складні (рис. 11). ДНК-геліказа, дві ДНК-полімерази на обох дочірніх ланцюгах і праймаза на відстаючому ланцюзі утворюють єдиний поліферментний комплекс – реплісому. Відстаючий ланцюг згинається таким чином, що її ДНК полімераза комплексує з ДНК-полімеразою лідируючого ланцюга. Цей вигин підводить 3'-кінець кожного вже синтезованого фрагмента Оказакі до тієї ділянки, де починається синтез нового фрагмента Оказакі. Комплекс геліказа-праймаза-полімераза рухається разом з вилкою реплікації і синтезує нові РНК-праймери. ДНК полімераза на відстаючому ланцюзі просувається вперед разом з репликаційною виделкою і багаторазово використовується для синтезу фрагментів Оказакі. Таким чином, ДНК синтезується однаково ефективно на обох материнських ланцюгах ДНК (матрицях). Ця схема реплікації ДНК отримала назву «моделі тромбона» (рис.11).
Рис. 11.- Схема реплікації ДНК у відповідності з «моделлю тромбона» [Жимульов І.Ф., 2002] Термінація. При двонаправленому синтезі реплікація закінчується в термінус-центрі (terminus C). Вилки, що рухаються назустріч, немов би збивають одна одну. При односпрямованій реплікації процес починається та закінчується в огі С. Знов утворена ДНК прикріплюється до виросту цитоплазматичної мембрани і, за рахунок локального синтезу мембрани дві молекули ДНК розводяться до полюсів клітини. Ці молекули можуть виявитися зчепленими, тобто утворюються катенани (синоним: катемери), що складаються з двох кільцевих молекул, зчеплених механічно. В їхньому від’єднанні беруть участь гіраза та топоізомераза І. Після утворення міжклітинної перетинки між двома молекулами ДНК формуються дві нові дочірні клітини. Процесинг. Відразу після реплікації у новоствореній ДНК в сайтах (ділянках) рестрикції відбувається метилювання аденіну, гуаніну та цитозіну. Це необхідно для мічення власної ДНК і запобігання розрізання ДНК в цих сайтах власними рестриктазами (ендонуклеазами рестрикції). Система рестрикції необхідна для спрямованого знищення чужорідної ДНК (наприклад, вірусної), проникаючої в клітину.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 1650; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.6.144 (0.009 с.) |