Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механізми реплікації у вірусів, прокаріот і еукаріотСодержание книги
Поиск на нашем сайте
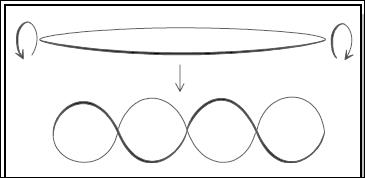
Репліка́ція (від лат. replicatio — відновлення) — процес синтезу дочірньої молекули дезоксирибонуклеїнової кислоти на матриці батьківської молекули ДНК. Під час наступного поділу материнської клітини кожна дочірня клітина одержує по одній копії молекули ДНК, котра є ідентичною ДНК вихідної материнської клітини. Цей процес забезпечує точну передачу генетичної інформації з одного покоління клітин іншому. Реплікацію ДНК здійснює складний ферментний комплекс, що складається з 15—20 різних білків, що називається реплісомою (англ. replisome). Для того щоб пояснити, яким чином може самокопіюватися (редуплікуватися) така стабільна та замкнута на собі структура, як подвійна спіраль ДНК, у 1953 році Уотсон і Крик припустили, що її ланцюги здатні до розкручування і подальшого часткового поділу внаслідок розриву водневих зв'язків у кожній комплементарній парі основ. Утворені одноланцюгові ділянки батьківської молекули можуть служити матрицею, до якої на основі комплементарності основ приєднуються відповідні нуклеотиди. Ці нуклеотиди з’єднуються між собою фосфодиефірними зв'язками з утворенням нового ланцюга, комплементарного батьківському. Оскільки цей процес відбувається на кожному з розділених ланцюгів вихідної молекули, то в результаті утворюються дві дволанцюгові структури, ідентичні батьківській ДНК. Такий механізм реплікації отримав назву напівконсервативного, оскільки в кожній з новоутворених молекул один ланцюг є старим (батьківським), а інший — знов синтезованим (дочірнім). Цей механізм забезпечує можливість такого розподілу ДНК між клітинами, що діляться, при якому кожна дочірня клітина отримує гібридну дволанцюгову молекулу ДНК, яка складається з батьківського та знов синтезованого ланцюгів. Перші дані на користь гіпотези напівконсервативного механізму синтезу ДНК були отримані Дж.Тейлором із співавторами (1957) цитологічним методом при вивченні реплікації хромосом кінських бобів (Vicia faba). Експериментально ця гіпотеза була доведена за допомогою фізико-хімічних методів М. Мезельсоном і Ф. Сталем (1958). Сутність їх досліду полягала в наступному. Бактерії (Е. coli) протягом багатьох генерацій вирощували в середовищі, яке містить в якості джерела азоту тільки важкий ізотоп N15. Ця мітка включалася в азотвмісні пуринові і пиримидинові основи, внаслідок чого ДНК в клітинах, вирощених у середовищі з N15, мала більш велику молекулярну масу на одиницю об'єму (тобто щільність), ніж ДНК в клітинах, вирощених у звичайних умовах, у присутності легкого ізотопу N14. Тому якщо клітини після тривалого вирощування на середовищі з N15 відмивали і переносили на час, що дорівнює одній, двом і т. д. генераціям, в середовище з N14 замість N15, то це повинно було привести до появи молекул ДНК з меншою щільністю. По закінченні однієї генерації клітин (протягом періоду, необхідного для їх подвоєння) після перенесення з середовища з N15 в середовище з N14 в клітинах з'явилася гібридна за щільністю ДНК, у якій один ланцюг був «важким», а інший — «легким». Якщо ДНК виділяли з клітини через дві генерації після такого перенесення, то частка «гібридної» ДНК становила лише половину всієї ДНК. У наступному поколінні ця «легка» фракція збільшувалася і становила 75% тотальної ДНК. Таку зміну можна пояснити тільки з позицій уявлення про напівконсервативний спосіб реплікації ДНК. На основі цих даних Мезельсон і Сталь прийшли до висновку, що в суворій відповідності з гіпотезою Уотсона-Крика кожний дочірній дуплекс ДНК після двох циклів подвоєння клітин містив один батьківський і один новостворений ланцюг ДНК. Такий механізм реплікації названий напівконсервативним. Отримані результати повністю виключили ймовірність консервативного способу реплікації, при якому одна дочірня ДНК мала б містити обидві вихідні ланцюги, а інша складалася б із двох новосинтезованих ланцюгів. Таким чином, в 1957 році Мезелсон і Сталь експериментально довели наявність у Е.coli напівконсервативного механізму реплікації. Вченимибули запропоновані щедва інших гіпотетичних механізми реплікації: консервативний і дисперсивний. Вони поширені значно рідше. При консервативному механізмі на одному з материнських ланцюгів здійснюється синтез дочірнього ланцюга, а утворений дочірній ланцюг служить матрицею для синтеза ще одного дочірнього ланцюга. Після поділу клітини в одну з новоутворених клітин попадає 2 старі ланцюги ДНК, а в іншу – 2 нові. Цей механізм випадковий і майже не зустрічається в природі, оскільки дає можливість накопичення мутацій у половині клітин популяції. Дослід Мезельсона і Сталя дозволив відкинути можливість дисперсивного механізму реплікації, при якому кожний дочірній ланцюг ДНК складається з коротких ділянок як батьківської, так і новоутвореної ДНК, сполучених між собою випадковим чином. Дисперсивний механізм припускає можливість рекомбінацій. Посля такої реплікації в дочірніх клітинах мають опинитися ДНК, що складаються з фрагментів старого та нового матеріалу (в еукаріот це відбувається в результаті кросинговера, а не реплікації). Сформульований Мезельсоном і Сталем висновок був повністю підтверджений і для інших об'єктів, включаючи тварин і вищі рослини. Разом з тим залишалося незрозумілим, в якому напрямку відбувається реплікація ДНК і які ферменти забезпечують цей процес. Відповідь на перше питання отримав Дж. Кернс (1963) в дослідах на Е. сoli. Подальший аналіз результатів досліду Кернса і дані інших експериментів, спрямованих на вивчення механізму реплікації ДНК у бактерій, фагів, плазмід, показали, що в більшості випадків вона відбувається двоспрямовано і починається, як правило, від одного унікального локусу ori. У цьому місці в одній з ланцюгів ДНК розривається фосфодиефірний зв'язок, що забезпечує подальше розкручування дуплексу та утворення особливих структур — репликативних вилок, що рухаються в протилежних напрямках по кільцевій ДНК (рис.3).
На електронномікроскопічних фотографіях молекула ДНК, що реплікується, нагадує своєю формою грецьку букву Ө (тета-тип); він характерний для прокаріот. Після того, як молекула ДНК буде реплікована повністю, обидві кільцеві копії опиняються «протягнутими» одна в іншу, як два кільця ланцюга. Вони мають бути роз’єднані. Цей процес здійснює топоізомераза II. Фермент розрізає одну з кільцевих молекул, скрізь розрив виводить петлю нерозірваної молекули, а потім з’єднує кінці розрізаної молекули, знов перетворюючи її в кільце. Розрізняють три основних типи реплікації: Ϋ-тип (ігрек), θ-тип (тета), σ-тип (сигма). Ϋ-тип реплікації характерний для лінійних хромосом еукаріотів та деяких вірусів, оскільки в них реплікативна вилка нагадує букву "ігрек". Реплікація кільцевих хромосом бактерій, плазмід, фага λ супроводжується локальним розходженням ланцюгів кільцевої ДНК в oriC і нагадує грецьку букву «тета», тому і отримала назву θ-типу. σ-тип реплікації характерний для вірусів з одноланцюговою ДНК чи РНК і нагадує кільце, що котиться або рулон, що розгортається. При цьому матрична нуклеїнова кислота нагадує кільце, що обертається, навколо якого синтезується дочірній ланцюг. Причому сформований дочірній ланцюжок може містити кілька тандемно розташованих копій генома вірусу, які згодом "розрізаються" рестриктазою. В еукаріотів реплікація відбувається перед поділом клітини, у прокаріотів — протягом всього життєвого циклу. У ДНК-містких вірусів, бактерій, ціанобактерій, а також в еукаріотичних органелах, що самоподвоюються (пластидах, мітохондріях, центріолях), хромосома уявляє собою двоспіральну, позбавлену гістонів, молекулу ДНК у вигляді кільця, закрученого в шпильку, завдяки чому хромосома має суперспіралізований вигляд (рис.4).
Рис.4. - Кільцева та суперспіралізована форма молекули ДНК у бактерій (за Жимульовим, 1998) Реплікація цих хромосом починається з однієї єдиної точки ініціації реплікації (т.і.р.) (рис.2) та прогресує до закінчення реплікації всієї хромосоми; отже, хромосома уявляє собою одну одиницю реплікації – реплікон. У більшості випадків (у бактерій – завжди) реплікація хромосоми відбувається в обидва боки від точки ініціації реплікації (ϴ-тип) (рис.5).
Рис.5.- Схема реплікації кільцевої ДНК - ϴ-тип (тета-тип) Процес реплікації прокаріот завершується утворенням двох дочерніх молекул ДНК. Їхні нуклеотидні послідовності ідентичні нуклеотидним послідовностям материнської подвійної спіралі ДНК. Завершення реплікації відбувається в спеціальних ділянках ДНК. В E. coli вони називаються ter -сайтами та містять коротку послідовність (близько 23 пар) нуклеотидів (рис.6).
Рис. 6. - Дві групи термінації реплікації у E. coli [Жимульов І.Ф., 2002] В E. coli знайдені дві групи ter -сайтів (рис. 6). У першій групі ter -сайтів зупиняється рух першої реплікаційної вилки, в другій групі ter -сайтів – рух другої. Вони розміщуються приблизно в 100 тпн від тієї точки, в котрій зустрічаються вилки реплікації. Для закінчення реплікації ДНК необхідний спеціальний білок. У бактерії E. coli він кодується геном tus. Цей білок розпізнає термінаторну послідовність, зв’язується з нею та зупиняє подальше просування вилки реплікації. Другий шлях, за яким може здійснюватися реплікація дволанцюгових кільцевих молекул ДНК, отримав назву «кільця, що котиться» («rolling circle») - σ – тип (сигма-тип); так реплікуються кільцеві ДНК деяких фагів, вірусів, мітохондрій, плазмід (рис.7). Таким способом утворюється багато нових молекул вірусних ДНК, а також здійснюється статевий процес у бактерій і ампліфікація генів. Реплікація ДНК за типом кільця, що котиться, ініціюється спеціальним ферментом, який кодується ДНК плазміди або бактеріофага; цей фермент розриває один із ланцюгів двохниткової кільцевої молекули в сайті DSO (англ. double-strand origin) (рис.) з утворенням «+»-ланцюга, що призводить до появи однониткових 5'- і 3'-кінців. Ініціаторний білок зв’язується з 5'-фосфатним кінцем розірваного ланцюга, а вільна 3'-OH група служить праймером для синтеза другого ланцюга ДНК-полімеразою III. 5'-кінець витісняється з подвійної спіралі та приєднується до клітинної мембрани; з ним зв'язується фермент ДНК-полімераза ІІІ. По мірі розкручування молекули величина звільненої одноланцюгової матриці від кінця 5' збільшується. На ній синтезується комплементарна дочірня нитка ДНК. У результаті утворюється та поступово подовжується вільний «хвіст». Використовуючи нерозірваний ланцюг як шаблон, реплікується вся кільцева молекула ДНК, заміщуючи розірвану одно ланцюгову ДНК за допомогою кодуємої реціпієнтом хелікази PcrA в присутності фактора ініціації реплікації. Синтез на кільцевій нитці може йти протягом декількох обертів молекули. В результаті довжина утвореної лінійної молекули може перевищувати довжину одного генома. Багаторазово повторений синтез копії ланцюга на кільцевій нитці ДНК призводить до появи лінійної нитки, яка містить кілька повторів того ж самого гена. Таку нитку називають конкатемером. Вона розрізається рестриктазами на частки, довжина яких дорівнює довжині хромосоми (геному). Після появи таких часток вони замикаються в кільцеві хромосоми. Здійснюється такий спосіб реплікації й у випадку переносу плазмідної або хромосомної ДНК від донора до реципієнта в процесі кон'югації у бактерій. Лінійні копії можуть перетворитися в кільцеву молекулу наступним чином. Спочатку ініціаторний білок робить розрив, термінуючи тим самим синтез іншого (провідного) ланцюга. Потім РНК-полімераза і ДНК-полімераза III реплікують одноланцюгову SSO-ДНК (англ. single-stranded origin), формуючи нове дволанцюгове кільце. Після цього ДНК-полімераза I замінює праймер на ДНК, а ДНК-лігаза зшиває кінці, утворюючи тим самим остаточну дволанцюгову ДНК. Механізм реплікації за типом кільця, що котиться, успішно використаний для ампліфікації ДНК з дуже невеликих кількостей ДНК вихідного зразка.
Рис.7. – Схема реплікації ДНК вірусного, бактеріального та еукаріотичного геномів У вірусів, що мають одноланцюгову ДНК, механізм її реплікації той самий, що і в дволанцюгових молекул, проте генетичну значимість в цьому випадку має тільки одна нитка. Цю нитку ДНК позначають знаком +. Коли (+)-нитка надходить у клітину, вона добудовує комплементарну нитку, утворюючи дволанцюгову молекулу ДНК, що містить (+)-нитку та (–)-нитку. Після розмноження вірусів в утворену частку фага включається тільки (+) нитка ДНК. Отже, реплікація може відбуватися або в одному, або в двох напрямках. При односпрямованій реплікації (характерна для еукаріот) уздовж ДНК рухається одна репликаційна вилка. При двобічній реплікації (ДНК бактеріальної хромосоми, вірусів, плазмід, що мають кільцеву форму) від точки початку в протилежних напрямках розходяться дві репликативні вилки. Результати дослідів показали, що розкручування двох комплементарних ланцюгів батьківської ДНК і їх напівконсервативна реплікація відбуваються практично одночасно і починаються в загальній точці початку реплікації, що позначається як локус ori (від англ. origin — початок). В еукаріот ініціація реплікації також відбувається в локусі ori та починається з появи Y-образної структури — реплікативної вилки (У-тип реплікації (ігрек-тип)), утворення якої пов'язане з розкручуванням дуплексу ДНК і локальним поділом її ланцюгів (рис.7). Оскільки два ланцюги дуплексу ДНК антипаралельні, синтез комплементарних ним ниток повинен в одному випадку йти в напрямку 5'-3', а в іншому — в напрямку 3'-5'. Перша нитка називається лідируючою, друга — запізнілою. Всі відомі ДНК-полімерази потребують для своєї активності вільного 3'-ОН кінця, до якого вони приєднують нуклеотиди. У результаті відбувається ріст ланцюга ДНК в напрямку 5'-3'. Як у такому випадку здійснюється синтез ланцюга ДНК в напрямку 3'-5'? Передбачалося, що для цього потрібна якась особлива ДНК-полімераза. Виявилося, що синтез 3'-5'-ланцюгів має перервчастий характер. Слід, однак, відзначити, що в ДНК еукаріотів, як правило, виявляється не один, а безліч локусів ori, що, мабуть, служить необхідною умовою для того, щоб величезні молекули ДНК в хромосомі еукаріотів встигли повністю відреплікуватися за час одного клітинного циклу. Поділ клітин еукаріотів відбувається шляхом мітозу, при цьому, щоб уникнути втрати генетичної інформації, спочатку подвоюється весь ядерний геном в S-фазі клітинного циклу. Тривалість S-фази становить 8 годин. ДНК центромер хромосом реплікується під час середньої стадії мітозу, що передує процесу сегрегації хромосом. Реплікація мітохондріальної та ядерної ДНК відбувається в різні фази клітинного циклу. Незважаючи на те, що загальна послідовність стадій при реплікації ДНК в еукаріотів і прокаріотів однакова, сам процес має незначні відмінності. Так, у еукаріотів під час реплікації ДНК (ядерна) залишається в нуклеосомній конфігурації. Фрагменти ДНК, багаті парами основ Г—Ц (R-смуги еухроматина в ущільненому хроматині), експресують гени «домашнього господарства», які функціонують у всіх клітинах організму. Ці фрагменти реплікуються на ранній стадії S-фази. Ділянки гетерохроматину, багаті парами основ А—Т (G-смуги), експресують невелику кількість генів і реплікуються на пізній стадії S-фази. Гени з великим вмістом пар А—Т, що кодують різні властивості і функціонують лише в певних клітинах, входять до складу факультативного гетерохроматину; їхня реплікація відбувається на ранній стадії S-фази тільки в тих клітинах, в яких вони експресуються, і на пізніх стадіях — в клітинах, де експресії не відбувається. Спостереження за допомогою електронного мікроскопу показали, що реплікація починається не з кінця лінійної молекули. Наприклад, у фага Т7, що має лінійну молекулу ДНК, реплікація починається на внутрішньому ділянці на 17% довжини всієї молекули, від лівого кінця.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 1443; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.148.115.187 (0.008 с.) |

 Рис.3.- Спрощена схема двоспрямованої реплікації кільцевих молекул ДНК бактерій при збереженні їх кільцевої структури
Рис.3.- Спрощена схема двоспрямованої реплікації кільцевих молекул ДНК бактерій при збереженні їх кільцевої структури