
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Адаптивные модификации, морфозы, фенокопии. В чем заключаются их различия?Содержание книги
Поиск на нашем сайте
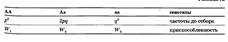
Адаптивные модификации, морфозы, фенокопии. В чем заключаются их различия? Как изменится частота аллелей а и А, при неодинаковой исходной приспособленности гомо- и гетерозигот в популяции приспособленности характеризуют динамику эволюционных изменений популяций, постоянно обсуждается в литературе. Дело в том, что изменения численностей и (или) частот в двух смежных поколениях недостаточны для характеристики направления эволюции популяции прежде всего потому, что сильно зависят и от случайных факторов, и от того, в какой фазе колебаний численности находится рассматриваемая популяция. Поэтому в качестве характеристики приспособленности предлагали использовать просто численность N. или для средней приспособленности N за ряд поколений.Рассмотрим действие отбора на популяцию, отвечающую всем условиям уравнения Харди-Вайнберга, за исключением одного: приспособленности разных генотипов различны. Предположим, также, что отбор и размножение разнесены во времени, скажем, отбираются неполовозрелые особи. Тогда, если частоты генов до отбора равны р и д, частоты генотипов, возникающих из гамет А и а, и их приспособленности соответствуют:
Следовательно, p2Wx, 2pqW2, q2W3. Это уже не частоты генотипов: их сумма равна не 1, а. Чтобы получить частоты генов после отбора, следует вышеприведенные выражения разделить на W
6.Как объяснить, что случайные мутации служат материалом направленного преобразования признаков? Мутагенез не направлен потому, что мутации сами по себе не повышают относительную приспособленность мутантных организмов к условиям их существования. Обычно они понижают приспособленность, но каждая — по своему. Однако, и мутационный процесс имеет ограничения. мутируют только те гены, которые имеются в генотипе. Комбинативная изменчивость уже не случайна. Она контролируется естественным отбором. Мутационная теория эволюции в первоначальном виде утратила свое значение тогда, когда выяснилось, что мутации возникают часто, а рецессивные мутации накапливаются в генофонде популяции, являясь основой генетической комбинаторики. регуляции морфогенеза еще не выявлена, очевидно, что мутации могут повлиять на него либо ускорив, либо замедлив, либо исказив формирование тех или иных компонентов формообразовательного аппарата, что влечет за собой более или менее сильное нарушение морфогенетических корреляций. Только сильные изменения начальных условий развития могут перевести морфогенез в другой креод, то есть исказить его или сделать дальнейшее развитие невозможным. Подобные изменения могут быть в той или иной степени компенсированы, благодаря существованию обратных связей между креодами, или в результате действия эргонтических корреляций. Однако, при отсутствии компенсации, они понижают приспособленность организма, в пределе до летали.
Каково значение для эволюции популяций мобилизационного резерва изменчивости. Может ли мутационный процесс сам по себе обеспечивать эволюцию? Объясните, почему большинство мутантных аллелей рецессивно по отношению к аллелям дикого типа? Что такое фенокопии? Возможный механизм их формирования. Примеры.
Фенокопии — изменения фенотипа под влиянием неблагоприятных факторов среды, по проявлению похожие на мутации. В медицине фенокопии — ненаследственные болезни, сходные с наследственными. Распространенная причина фенокопий у млекопитающих — действие на беременных тератогенов различной природы, нарушающих эмбриональное развитие плода (генотип его при этом не затрагивается). При фенокопиях изменённый под действием внешних факторов признак копирует признаки другого генотипа (пример - приём алкоголя во время беременности приводит к комплексу нарушений, которые до некоторой степени могут копировать симптомы болезни Дауна).
Механизмы возникновения мутаций и фенокопий различны: мутация является следствием изменения структуры гена, а фенокопия — результатом нарушения реализации наследственной информации. Фенокопии могут возникать и вследствие подавления функции определенных генов. В отличие от мутаций они не наследуются. Причинами фенокопий являются: 1. Кислородное голодание плода. 2. Болезнь матери при беременности. 3. Психическая травма у беременной. 4. Эндокринные заболевания у беременной 5. Питание беременной (недостатки С, В, Р, РР вит., Со, Са, Fe). 6. Лекарственные препараты при беременности (антибиотики, сульфаниламиды). К генетическим механизмам подавления действия аллелей можно отнести эпистаз. Это такой тип взаимодействия разных генов, при котором аллели одного гена подавляют (эпистатируют) действие другого. Эпистаз может быть доминантным, т. е. эпистатируют доминантные аллели, и рецессивный, когда эпистатируют рецессивные аллели. При длительном скрещивании расщепление у гибридов второго поколения изменяется с менделевского 3:3:3:1 при доминантном зпистазе на 12:3:1, или на 9:3:4 при рецесивном. Понимание механизма эпистаза кроется в биохимических процессах: при многоэтапном процессе биосинтеза продукта, участвующего в формировании анализируемого признака, ген, включающийся в работу раньше, может эпистатировать более "поздний" ген. Примеры реакция человеческих эмбрионов на медицинский препарат талидомид – ненормальное развитие рук. Наблюдались нехватка пальцев, недоразвитие и даже полное отсутствие рук. во время дробления можно получить фенотип, имитирующий мутацию tetraptera—четырехкрылости (рис. 23). Реакция на эти воздействия среды: и талидомид для людей, и эфирный наркоз или тепловой шок для дрозофилы не адаптивные реакции организма. Тот факт, что морфоз может копировать фенотипическое проявление мутации свидетельствует о том, что и внешние (средовые) и внутриорганизменные (генетические) повреждающие факторы нарушают одни и те же формообразовательные процессы.
28. Эволюционное значение "принципа основателя" и "эффекта бутылочного горлышка". Принцип основателя - биологический закон, согласно которому особь-основатель новой изолированной колонии или островной популяции несет в себе лишь незначительную часть генетической информации, заложенной в популяции или виде, откуда происходит особь-основатель. Аллопатрическое видообразование — широко распространенный способ возникновения новых видов. При расширении ареала окраинные популяции — краевые изоляты — хорошо изолированы от популяций, обитающих в более благоприятных частях ареала, а численность особей, составляющих такой изолят невелика. В краевых изолятах возникает эффект горлышка бутылки—генофонд их обеднен. В то же время, популяции, осваивающие новые территории, вынуждены приспосабливаться к новым условиям среды. Это создает новые векторы отбора. Сочетание изменений материала для отбора, изоляции и изменения направления отбора обеспечивает дивергенцию. Эта схема, предложенная Майром, уточнена введенным им «принципом основателя». Немногочисленные особи — основатели краевых изолятов несут генофонд, отличающийся по частотам аллелей от генофонда других популяций того же (исходного) вида, что создает стартовые условия быстрой дивергенции.
Имеются данные и подтверждающие существование принципа основателя, и противоречащие ему. Формозов и Яхонтов (1988) показали, что у пищух Ochotona alpina—hyperboreum complex, обитающих на северо-востоке Якутии, звуковые сигналы мономорфны, что отличает эти популяции от полиморфных по вокализации популяций пищух плато Путорана. Авторы объясняют стабилизацию звукового сигнала в краевых изолятах принципом основателя.. С другой стороны, Шилов и др. изучали береговые и островные популяции полевки-экономки Microtus oeconomus в Дарвинском заповеднике. Островные популяции были изолированы от береговых с момента заполнения Рыбинского водохранилища, в течение, минимум, 40 поколений. Несмотря на сильные различия экологических условий на материке и на торфяных островах, всплывших вместе с полевками при затоплении водохранилища и, затем, закрепившихся на отмелях корнями макрофитов, различий морфофизиологических показателей не было обнаружено. Экологические же различия велики. Количество примеров и подтверждающих, и не подтверждающих принцип основателя может быть увеличено. Их совокупность позволяет считать этот принцип не универсальным пусковым механизмом аллопатрического видообразования.
Что такое атавизм? Атавизмом называется восстановление у потомков признаков, свойственных отдаленным предкам и утраченных предками более близкими. Атавизмы всегда проявляются как индивидуальные изменения. Этим они отличаются от рудиментов. Различают гибридный атавизм и атавизм процессов онтогенеза. Гибридный атавизм чаще всего проявляется у домашних животных при межпородных или межлинейных скрещиваниях восстановлением окраски, приближая ее к дикому типу. Впервые это явление описал Дарвин у домашних голубей. Часто оно встречается при скрещивании разных пород кроликов и разных линий лабораторных мышей. Гибридный атавизм объясняется тем, что окраска домашних животных обычно обусловлена рецессивными аллелями. При скрещивании разных пород или линий увеличивается гетерозиготность, что ослабляет фенотипический эффект рецессивов и восстанавливает исходную окраску. Атавизмы, возникающие спонтанно, в результате восстановления свойственных предкам процессов морфогенеза или прй регенерации, можно, по-видимому, объяснить тем, что редукция не всегда связана с полной утратой морфогенетических корреляций, определявших развитие признака у предков. Поэтому, мутационное или средовое нарушение морфогенеза потомков может, в той или иной степени, восстановить утраченные мор- фогенетические зависимости и привести к возникновению атавизмов. Таковы атавизмы трехпалости у домашней лошади, полимастии или возникновение хвоста у людей. Подобными же причинами объясняется сходство регенерировавшей клешни речного рака Astacus fluviatilis с нормальной клешней более примитивного вида A. leptodactilus.
Некогерентной? Адаптивной зоной (АЗ) называется совокупность условий, в пределах которых развивается данный таксон. В процессе развития популяций различных организмов в пределах одной экологической ниши, происходит постепенное распределение пространства и ресурсов между организмами. В результате этого каждый приспосабливается к конкретной АЗ. Происходит специализация организмов. В результате этого попаданию вида в другую АЗ препятствует мощная конкуренция. Подобное ограничение способствует дальнейшей специализации в использовании «своего» ресурса. В результате таких взаимодействий (реализующихся в основном в коадаптации видов) происходит медленная направленная эволюция видов – когерентная эволюция (КЭ). КЭ основана на тесном взаимодействии и согласовании элементов сложной системы (экологической ниши). КЭ происходит значительно более медленными темпами, чем некогерентная. Также при когерентной эволюции вероятность становления новых жизненных форм значительно ниже, чем при некогерентной. Возникновение некогерентной фазы эволюции часто связано с экологическими кризисами, в результате чего взаимосвязи между элементами системы (видами) разрушаются, степень синхронизованности в развитии между разными группами падает. В результате различные виды обособляются в виде группировок. Подобное разрушение целостной (когерентной) структуры дает возможность более интенсивному проникновению в систему новых видов. Характерные времена некогерентной эволюции значительно выше, чем у когерентной.
Что такое педоморфоз? Педоморфоз (неотения) – эволюционное преобразование организмов, характеризующееся полной утратой взрослой стадии и соответствующим укорочением онтогенеза, в котором последней становится стадия, бывшая прежде личиночной. Педоморфоз возможен у тех видов организмов, личинки которых приобрели способность к размножению, в частности, на основе неотении. Путём педоморфоза возникли некоторые группы хвостатых земноводных (протеи, сирены).
Что такое ценогенез? Ценогенез первоначально был выделен Северцовым как один из способов достижения биологического прогресса. Ценогенез – выработка провизорных приспособлений, обеспечивающих выживание организма на ранних стадиях онтогенеза. Шмальгаузен вывел это понятие из способов достижения биологического прогресса, так как оно содержится (аналогично) в остальных (например ароморфозу или идиоадаптации). Пример: появление вторичных яйцевых оболочек у амниот – типичный ароморфоз. Развитие яйцевого зуба для вскрытия скорлупы у детенышей крокодилов или птенцов птиц – идиоадаптация.
Адаптивные модификации, морфозы, фенокопии. В чем заключаются их различия?
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-04-23; просмотров: 1307; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.0.34 (0.009 с.) |

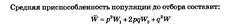 Вклады прошедших отбор генотипов в следующее поколение будут,
Вклады прошедших отбор генотипов в следующее поколение будут,

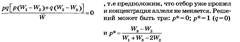 Два первых решения соответствуют полному вытеснению одного аллеля другим, их принято называть тривиальными. Третье, не тривиальное, решение существует при следующих условиях: 0<р<1, это значит, что W3-W2>0, W1+W3-2W2>0, W1 + W3-2W2>W3-W2 (эти условия выполняются при W3>W2 и W1>W2); или W3-W2<0, W^+Wg-W2<0, Wx+W3 - 2W2 < W3 - W2 (эти условия выполняются при W3 <W2 и W2 >WX). Таким образом, нетривиальное решение возможно лишь в тех случаях, когда приспособленность гетерозиготы либо выше приспособленности обеих гомозигот, либо ниже их. При равенстве приспособленностей гомозигот и гетерозиготы нетривиального решения не существует.
Два первых решения соответствуют полному вытеснению одного аллеля другим, их принято называть тривиальными. Третье, не тривиальное, решение существует при следующих условиях: 0<р<1, это значит, что W3-W2>0, W1+W3-2W2>0, W1 + W3-2W2>W3-W2 (эти условия выполняются при W3>W2 и W1>W2); или W3-W2<0, W^+Wg-W2<0, Wx+W3 - 2W2 < W3 - W2 (эти условия выполняются при W3 <W2 и W2 >WX). Таким образом, нетривиальное решение возможно лишь в тех случаях, когда приспособленность гетерозиготы либо выше приспособленности обеих гомозигот, либо ниже их. При равенстве приспособленностей гомозигот и гетерозиготы нетривиального решения не существует.


