Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Топическая диагностика заболеваний нервной системыСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
ПРОВОДНИКИ ЧУВСТВИТЕЛЬНОСТИ
Клетки первых, или периферических, чувствительных нейронов заложены в межпозвоночных спинальных ганглиях; отростки их — чувствительные волокна периферических нервов — проводят импульс с периферии от нервных чувствительных окончаний.
Рис. 1. Поперечный срез спинного мозга. [Формирование центростремительных проводников из волокон задних чувствительных корешков и подход волокон центробежных путей к клеткам передних рогов. 1 — волокна суставно-мышечной, вибрационной и тактильной чувствительности; 2 — волокна болевой и температурной чувствительности; 3 — волокна мозжечковых проприоцепторов; 4 — дорсальный спино-церебеллярный путь; 5 — вентральный спино-церебеллярный путь; 6 — пучок Бурдаха; 7 — пучок Голля; 8 — спино-таламический путь.
От клеток межпозвоночных узлов импульс по волокнам заднего чувствительного корешка проводится в спинной мозг, где волокна различных видов чувствительности расходятся в разных направлениях (рис. 1). Проводники суставно-мышечного чувства, вибрационного и части тактильного, не заходя в серое вещество спинного мозга, вступают прямо в задний столб своей стороны и в составе пучков Голля и Бурдаха поднимаются, не прерываясь, вверх до продолговатого мозга, где и заканчиваются в ядрах задних столбов или ядрах Голля и Бурдаха (рис. 2). Следовательно, первые, или периферические, нейроны названных видов чувствительности обеспечивают проведение импульса от периферии до продолговатого мозга, причем проводники следуют все время по своей стороне спинного мозга.
Рис. 2. Ход чувствительных путей.
Так как проводники суставно-мышечного чувства, тактильного и других вступают в задние столбы с их наружной стороны, рядом с задним рогом, то происходит постепенное оттеснение ранее шедших в заднем столбе волокон (от нижележащих сегментов) к средней линии. В силу этого в медиально расположенном пучке Голля оказываются проводники от нижних конечностей, в латеральном же пучке Бурдаха — проводники от более высоких сегментов, главным образом верхних конечностей. Проводники болевого и температурного чувства, а также некоторой части тактильного, вступают из заднего корешка в задний рог спинного мозга, где аксоны первого, или периферического, нейрона заканчиваются, контактируя с клетками, здесь расположенными (второй нейрон). Аксоны этих клеток переходят через переднюю серую спайку на противоположную сторону (см. рис. 1), делая здесь перекрест, и вступают в боковой столб, составляя tractus spino-thalamicus. Названный проводник составляется, следовательно, из волокон вторых нейронов болевого и температурного чувства противоположной стороны (частью и тактильного). Переход перекрещивающихся волокон происходит не в горизонтальной плоскости, на уровне данного сегмента, а косо — вверх, в результате чего вхождение волокон данного сегмента в tractus spino-thalamicus осуществляется на 2 — 3 сегмента выше. Это определяет соответствующее снижение верхнего уровня проводниковых расстройств болевого и температурного чувства на противоположной стороне при поражении бокового столба. Не прерываясь, tractus spino-thalamicus проходит по боковому столбу спинного мозга в продолговатый, в варолиев мост и ножки мозга, оканчиваясь в латеральном ядре зрительного бугра. Волокна от клеток заднего рога вступают в tractus spino-thalamicus изнутри, оттесняя тем самым волокна, идущие в пучке от нижележащих сегментов кнаружи. В итоге в спино-таламическом тракте волокна располагаются таким образом, что в наружном его отделе проводится чувствительность от нижних сегментов (крестцовых, поясничных), во внутреннем же — от верхних (так называемый закон эксцентрического расположения более длинных путей). Указанное обстоятельство имеет значение для топической диагностики заболеваний спинного мозга: при процессах интрамедуллярных, исходящих из серого вещества в направлении бокового столба, проводниковые расстройства болевого и температурного чувства, начинаясь с уровня пораженных сегментов, спускаются по мере развития процесса вниз. При процессах же экстрамедуллярных, воздействующих на tractus spino-thalamicus снаружи, проводниковые расстройства нарастают снизу вверх. Вместе с волокнами болевого и температурного чувства из заднего корешка переходят в задний рог и волокна проприоцепторов мозжечка. Периферические нейроны заканчиваются в сером веществе спинного мозга (см. рис. 1). Здесь заложены клетки вторых нейронов: аксоны их выходят в боковой столб своей стороны и располагаются здесь на периферии, составляя tractus spino-cerebellaris dorsalis (пучок Флексига) и ventralis (пучок Говерса). Названные проводники поднимаются по спинному мозгу вверх и заканчиваются в черве мозжечка. Подробнее о спино-церебеллярных путях будет изложено ниже (глава IV). Вернемся к рассмотрению хода путей суставно-мышечного и тактильного чувства. Как было указано выше, волокна первых нейронов закончились в продолговатом мозге, в ядрах Голля и Бурдаха. Отсюда, от клеток названных ядер, аксоны вторых нейронов направляются вентрально и к средней линии, совершая на уровне олив перекрест (в межоливном слое). Переходя на противоположную сторону, названные волокна (fibrae arcuatae internae) присоединяются, прилегая изнутри, к tractus spino-thalamicus (см. рис. 2). Пучок волокон вторых нейронов суставно-мышечной и тактильной чувствительности носит название tractus bulbo-thalamicus. Слияние обоих чувствительных путей — tractus spino-thalamicus и bulbo-thalamicus, начавшись в продолговатом мозге, завершается окончательно только в мосту. Общий чувствительный путь мозгового ствола, составленный двумя названными пучками, носит наименование медиальной петли — lemniscus medialis (в классических описаниях медиальной петлей именуется только tractus bulbo-thalamicus). Медиальная петля представляет собой собрание волокон вторых нейронов всех видов чувствительности противоположной стороны тела (перекрест волокон болевой и температурной чувствительности происходит последовательно по сегментам на всем протяжении спинного мозга в передней серой спайке, а волокон суставно-мышечного и тактильного чувства — в межоливном слое продолговатого мозга). Медиальная петля располагается в среднем этаже ствола: в продолговатом мозге и мосту — над пирамидами, в ножках мозга — над substantia nigra. Находясь вначале вблизи от средней линии, обе медиальные петли в мосту начинают расходиться, располагаясь постепенно все более латерально. К петлям присоединяются волокна от ядер чувствительных черепных нервов: языко-глоточного, блуждающего и тройничного (тоже после перекреста). Волокна медиальной петли (tractus spino-thalamicus и tractus bulbo-thalamicus) заканчиваются в латеральном ядре зрительного бугра. Часть волокон заканчивается и в медиальном ядре; они, по-видимому, устанавливают рефлекторные связи в пределах подбугровых и подкорковых образований. В зрительном бугре заложены клетки третьих нейронов чувствительности, волокна которых составляют tractus thalamo-corticales, направляющиеся через внутреннюю капсулу (задняя треть заднего бедра — см. рис. 55, VII) и corona radiata в кору головного мозга, в заднюю центральную извилину и теменную долю. Проекция в кору рецепторных полей противоположной стороны тела осуществляется следующим образом: в верхнем отделе задней центральной извилины представлены рецепторы ноги, в среднем — руки и в нижнем отделе — головы, т. е. в порядке, обратном расположению частей тела. Анализ и синтез ощущений как от кожных рецепторов (экстероцепторы), так и су-ставно-мышечных (проприоцепторы), происходит не только в задней центральной извилине, но и в значительно более широких территориях коры, в частности, в теменной доле, причем в последней в основном представлена глубокая чувствительность.
ИЗМЕНЕНИЯ РЕФЛЕКСОВ
Рефлексы могут изменяться в сторону: 1) понижения или утраты их, 2) повышения и 3) извращения. Наконец, в патологических условиях могут появляться новые рефлексы, в нор мене вызываемые. Утрата или понижение рефлексов (арефлексия и гипорефлексия) возникает в результате нарушения целости и проводимости рефлекторной дуги в любом ее отделе (афферентном, сочетательном, эфферентном). Симметричное понижение и даже утрата рефлексов иногда не является еще признаком поражения нервной системы. Так, некоторые рефлексы у известного числа здоровых лиц вызываются с трудом или не получаются вовсе. Чаще это относится к глубоким рефлексам верхних конечностей (сухожильные рефлексы нижних конечностей отличаются большим постоянством, и их отсутствие всегда требует тщательного и всестороннего исследования нервной системы). Брюшные рефлексы иногда не могут быть вызваны при дряблой брюшной стенке. Брюшные рефлексы, как кожные, так и глубокие, понижаются или исчезают не только при поражении их рефлекторных дуг, но и при разобщении соответствующих им спинномозговых сегментов (DVII — DХII) с корой головного мозга, т. е. при поражении пирамидных путей. Объясняется это тем, что брюшные рефлексы не являются врожденными, а вырабатываются в связи с развитием коры головного мозга и пирамидной системы в течение индивидуальной жизни. Появление их совпадает с временем выработки у ребенка вертикального положения тела. Брюшные рефлексы, таким образом, стимулируются корой головного мозга и поэтому исчезают при разобщении с ней. Понятно, что они угасают также и при перерыве их рефлекторных спинальных дуг. Повышение рефлексов (гиперрефлексия) указывает на усиление рефлекторной деятельности сегментарного аппарата (спинного мозга, мозгового ствола). Чаще всего причиной повышения рефлексов является поражение пирамидных путей — системы через которую передаются тормозящие влияния коры головного мозга на рефлекторные сегментарные спинномозговые механизмы. Опять-таки симметричное повышение рефлексов при отсутствии других патологических симптомов не всегда говорит о наличии органического заболевания; высокие рефлексы могут быть и у здоровых лиц, могут наблюдаться у невротиков, при некоторых интоксикациях и т.д. Крайним проявлением повышения сухожильных рефлексов являются так называемые клонусы. Клонусы представляют собой ритмические сокращения какой-либо мышцы, возникающие в результате растяжения ее сухожилия. В сущности, клонус есть цепь следующих один за другим сухожильных рефлексов, вызываемых непрерывающимся растяжением сухожилия. Наиболее частыми являются клонусы коленной чашки и стопы. Клонус коленной чашки вызывается резким смещением коленной чашки книзу, причем оттянутая коленная чашечка продолжает удерживаться в смещенном положении. Исследуемый при этом лежит на спине с выпрямленными ногами. Коленная чашка захватывается большим и указательным пальцами исследующего и толчкообразно смещается вниз. Натягивается сухожилие m. quadricipitis, прикрепляющего мышцу к верхнему краю сумки коленной чашки, что при очень высоком коленном рефлексе бывает достаточным, чтобы вызвать сокращение мышцы; растяжение сухожилия не прекращается, и сокращения мышцы следуют одно за другим, обусловливая ритмическое движение коленной чашки. Клонус стопы вызывается также в лежачем положении исследуемого. Правой рукой стопа захватывается за ее дистальную часть, нога сгибается в коленном и тазобедренном суставах; резким толчком стопа разгибается в голеностопном суставе. В результате растяжения при этом ахиллова сухожилия возникают (при крайней степени живости ахиллова рефлекса) ритмические движения сгибания и разгибания стопы. Так как клонусы коленной чашки и стопы суть лишь показатели значительного повышения коленного и ахиллова рефлексов, то они могут наблюдаться во всех случаях гиперрефлексии, в том числе и не при органическом поражении нервной системы. В отличие от клонусов органических, клонусы при неврозах, физиологическом повышении рефлексов и т.д. обычно недостаточно стойки, всегда равномерно выражены с обеих сторон и не сопровождаются другими органическими симптомами. Клонусы на верхних конечностях наблюдаются редко; чаще других бывает клонус кисти, возникающий в результате резкого толчкообразного разгибания ее. Если симметричное понижение или повышение рефлексов не всегда является признаком поражения нервной системы, то неравномерность их всегда указывает на существующее органическое заболевание. Неравномерность рефлексов (анизорефлексия) возникает либо в результате понижения рефлексов на одной стороне (поражение рефлекторной дуги в нерве, корешках или сером веществе спинного мозга), либо повышения его на другой (поражение пирамидного пути). Установление неравномерности рефлексов является, таким образом, чрезвычайно важным. Поэтому исследование их должно производиться тщательно; удары молоточком, штриховые раздражения и т.д. должны наноситься точно и быть одинаковой силы при исследовании справа и слева; желательно не ограничиваться однократным исследованием, вызывать рефлексы разными методами и т.д. Патологические рефлексы. К этой группе относятся извращенные рефлексы или такие, которые в норме не существуют и вызываются лишь в патологических условиях, т. е. при поражении нервной системы. К числу патологических рефлексов, осуществляемых ротовой мускулатурой, относятся следующие. Назо-лабиальный рефлекс (М.И. Аствацатуров), вызываемый постукиванием молоточком по спинке носа. Ответная реакция заключается в сокращении m. orbicularis oris, круговой мышцы рта (вытягивание губ вперед). Хоботковый рефлекс — та же реакция, возникающая при ударе молоточком по верхней или нижней губе. Сосательный рефлекс получается в результате прикосновения к губам или штрихового раздражения их; в ответ наблюдаются сосательные движения губами. Дистанс-оральный рефлекс, клиническое значение которого подчеркивает С.И. Карчикян, вызывается не непосредственным раздражением губ, а лишь приближением ко рту больного молоточка: еще до удара им возникает «хоботковое» вытягивание губ вперед (сокращение m. orbicularis oris). Ладонно-подбородочный рефлекс (Маринеско — Радовичи) вызывается штриховыми раздражениями кожи ладони над thenaris, получается сокращение m. mentalis на данной стороне. т. е. нерезкое смещение кожи подбородка кверху. Перечисленные «оральные» рефлексы редко наблюдаются у здоровых взрослых людей; в норме они имеются у новорожденных и в младенческом возрасте. Появление их характерно для так называемого псевдобульбарного паралича, когда вследствие разобщения рефлекторных центров с корой головного мозга выпадают тормозящие влияния последней на сосательные автоматические реакции, связанные с сегментарными аппаратами ствола мозга. Они наблюдаются также в старческом возрасте. Группа оральных рефлексов относится к так называемым «аксиальным» (М.И. Аствацатуров) рефлексам, т. е. к тем видам двигательных реакций, которые вызываются с «оси» тела (голова, шея, туловище). Вместе с другими аксиальными оральные рефлексы могут быть повышены и при так называемом паркинсоновском синдроме, т. е. при экстрапирамидном парезе (гл. VII). На верхних конечностях в патологических условиях (при поражении пирамидных путей) может возникать ряд новых патологических рефлексов. Чаще других исследуется верхний симптом Россолимо: сгибание концевой фаланги большого пальца при коротком ударе пальцами исследующего по кончикам II — V пальцев исследуемого при свисающей свободно кисти. Модификацией методики является прием Е.Л. Вендеровича: при супинированной кисти исследуемого удар наносится по концам слегка согнутых II — V пальцев. Другие патологические рефлексы намечены слабо или не вызываются вовсе, почему большого распространения в клинике не получили. Упомянем только о рефлексе Клиппель — Вайля: при пассивном разгибании II — V пальцев кисти большой палец рефлекторно сгибается. Феномен наблюдается при центральном параличе или парезе верхней конечности (поражении пирамидных путей). В норме же происходит разгибание большого пальца. Основную и практически весьма важную группу патологических рефлексов составляют патологические рефлексы, обнаруживаемые на стопе. Кардинальными из них являются следующие. Симптом Бабинского — извращенный подошвенный рефлекс, или симптом разгибания большого пальца. В норме при штриховом раздражении подошвы получается рефлекторное сгибание всех пяти пальцев. При пирамидном поражении то же раздражение вызывает разгибание большого пальца, иногда изолированное, иногда с одновременным разведением остальных пальцев («знак веера»). Симптом весьма постоянный при центральных параличах и парезах; является одним из ранних и наиболее тонких проявлений нарушения целости пирамидного пути в головном или спинном мозге выше сегментов рефлекторной дуги подошвенного рефлекса (выше LV — SI сегментов). Симптом Россолимо — рефлекторное сгибание II — V пальцев стопы в результате короткого удара по кончикам названных пальцев пальцами исследующего или молоточком. Симптом Бехтерева — то же сгибание пальцев при постукивании молоточком по передне-наружной поверхности тыла стопы. Симптом Жуковского вызывается ударом молоточка по подошве под пальцами; ответным рефлекторным движением является подошвенное сгибание II — V пальцев. Симптом Оппенгейма получается в результате проведения с нажимом мякотью большого пальца исследующего по передней поверхности tibiae сверху вниз; феномен заключается в таком же разгибании большого пальца, как и при феномене Бабинского. Симптом Гордона — такое же рефлекторное разгибание большого пальца, но в результате сжатия рукой исследующего массы икроножных мышц. Симптом Шеффера — рефлекторное разгибание большого пальца при щипковом раздражении или сильном сдавлении ахиллова сухожилия. Симптом Гиршберга — сгибание и поворот стопы кнутри, вызываемые штриховым раздражением внутреннего края подошвы. Симптом Пуссепа — отведение V пальца при штриховом раздражении наружного края стопы. Описан как симптом экстрапирамидного пареза; по нашему мнению, является частным (неполным) проявлением знака веера, т. е. симптомом пирамидного поражения. Указанные здесь патологические рефлексы, обнаруживаемые на стопе, характерны для поражений пирамидной системы и представляют собой формы реакций нижележащего двигательного аппарата, разобщенного с корой головного мозга. В норме эти рефлексы наблюдаются у детей до того возраста, когда вырабатываются вертикальное положение тела и функция ходьбы, т. е. до 1 — 1,5 лет. В связи с развитием коры головного мозга старые типы реакций видоизменяются и могут выявиться вновь в прежнем их виде при освобождении нижележащих приборов от регулирующих и перестраивающих их деятельность (субординирующих) влияний коры головного мозга. Здесь приведены основные виды патологических стопных рефлексов, практически более ценные, имеющие наибольшее значение в клинической практике. Описано большое количество других патологических стопных феноменов: они отличаются от описанных обычно только местом и характером наносимого раздражения. Ответные реакции сводятся в основном к той же тыльной флексии большого пальца или сгибанию остальных пальцев. Защитные рефлексы также являются одним — из симптомов поражения пирамидного пути. Особенно отчетливо выражены они при поперечном поражении спинного мозга (разобщение нижележащих сегментов последнего от головного мозга). Вызываются лучше всего щипком, а также уколом (иногда только серией следующих один за другим уколов) или резким подошвенным сгибанием пальцев стопы, производимым исследователем (В.М. Бехтерев). Характеризуются довольно длительным скрытым периодом (замедленной реакцией); парализованная нижняя конечность при названных раздражениях «отдергивается», непроизвольно сгибаясь в тазобедренном, коленном и голеностопном суставах («реакция укорочения»); противоположная конечность (предварительно согнутая) при этом распрямляется, разгибаясь в суставах («реакция удлинения»). Поочередно нанося раздражения то на одну, то на другую ногу, можно получить имитацию «автоматизма» ходьбы. Защитные рефлексы могут иметь некоторое значение при определении уровня поперечного поражения спинного мозга: иногда они могут быть получены при раздражениях, наносимых на всю поверхность тела ниже расположения места перерыва спинного мозга (сдавления опухолью, травмы и др.). Следует учесть, что точные указания о нижней границе поперечного поражения спинного мозга на основании исследования защитных рефлексов, если они имеются, все же получить трудно. Тонические, или постуральные, рефлексы (рефлексы положения) представляют собой проявления особой сложной иннервационной системы, автоматически регулирующей тонус мышц для поддержания положения тела. Существуют механизмы, рефлекторно регулирующие равновесие как в покое, так и в движении. Перераспределение мышечного тонуса в значительной мере зависит от изменения положения в пространстве головы (лабиринтные и шейные тонические рефлексы). Как показал эксперимент на животных, основное значение в регуляции мышечного тонуса при стоянии и ходьбе имеют вестибулярные и красные ядра в мозговом стволе. В норме тонические рефлексы в значительной мере заторможены высшими отделами головного мозга и выявляются с особой интенсивностью при их выключении. Ослабление или усиление постуральных рефлексов при поражении тех или иных отделов нервной системы можно наблюдать и у человека, но далеко не с таким постоянством и закономерностью, как у экспериментальных животных. К числу тех симптомов, которые могут считаться проявлением расстройств постуральных рефлексов и которые отличаются несколько большим постоянством, относятся следующие. 1. Основной опыт. Исследуемый вытягивает руки вперед и с закрытыми глазами производит максимальный поворот головы вправо; обе руки при этом несколько отклоняются вправо же, причем правая рука, кроме того, еще и приподнимается несколько кверху. Соответствующее изменение положения рук происходит при последующем затем задании повернуть голову влево. Феномен усилен на стороне, где имеются мозжечковые расстройства, и ослаблен или утрачен при поражении экстрапирамидной системы. 2. Автоматическое приподнимание руки (больше правой) происходит через некоторое время, если исследуемому предложить держать руки вытянутыми вперед при закрытых глазах. Усиление поднимания происходит в случае мозжечкового поражения на соответствующей очагу стороне. 3. Пронационный феномен. При закрытых глазах руки вытягиваются вперед в положении полной супинации (ладонями кверху); постепенно возникает небольшая пронация. Усиление феномена наблюдается при пирамидных и мозжечковых поражениях. Не приводя других (весьма многочисленных) симптомов проявления постуральных рефлексов, укажем, что большого значения в клинике нервных заболеваний они не приобрели и широкого применения в исследовании нервной системы не получили.
ДВИЖЕНИЯ, ИХ РАССТРОЙСТВА
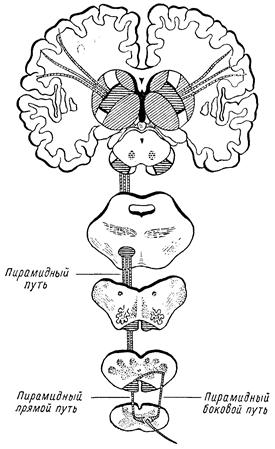
Двигательная функция человека представляется чрезвычайно сложной. В осуществлении движений имеет значение целый ряд отделов нервной системы. В одних случаях движения примитивны, происходят непроизвольно по типу простого рефлекторного акта и осуществляются за счет деятельности сегментарного аппарата (спинного мозга, мозгового ствола). Примером таких простых автоматических движений у человека являются разобранные выше рефлексы. Существуют, однако, и более сложные автоматизмы, осуществляемые не только сегментарным аппаратом, но и более высокими отделами нервной системы, в частности за счет экстрапирамидных иннервации. Ряд еще более сложных движений осуществляется не только за счет иннервации экстрапирамидной системы и сегментарного аппарата, но и при известном участии коры головного мозга, например ходьба в обычных условиях. Наконец, существует многочисленная категория движений, при которых участие коры головного мозга является основным, а иннервации экстрапирамидные и сегментарные — лишь подсобными. Сюда относится ряд целенаправленных действий, осуществляющих сложные акты двигательного поведения человека (праксии). Движения, возникающие в результате корковых иннервации, выработанные в жизненном опыте и являющиеся, по существу, условными рефлексами, обычно называются «произвольными»; автоматические рефлекторные движения могут быть названы «непроизвольными». Для осуществления «произвольного» движения необходимо, в частности, чтобы импульсы, возникшие в коре головного мозга, были проведены к мышце. Проведение импульса из коры происходит по цепи, состоящей из двух нейронов: 1) центрального двигательного и 2) периферического двигательного нейрона. Весь путь называется кортико-мускулярным (tractus cortico-muscularis). Центральный двигательный нейрон начинается от области коры, расположенной кпереди от роландовой борозды в передней центральной извилине, — в задних отделах верхней и средней лобных извилин и в lobulus paracentralis (см. рис. 65 и 67). Нервные клетки, дающие волокна для иннервации отдельных мышечных групп, имеют расположение, обратное расположению частей человеческого тела: проекция движений нижней конечности — в верхних отделах передней центральной извилины, верхней конечности — в среднем ее отделе, а головы, лица, языка, глотки и гортани — в нижнем. В заднем отделе верхней лобной извилины представлена проекция движений туловища и в заднем отделе средней лобной извилины — поворота головы и глаз в противоположную сторону. Иннервация мускулатуры перекрестная, т. е. правое полушарие связано с мускулатурой левой половины тела, а левое — с правой (перекрест проводников, о чем будет сказано ниже). Центральный двигательный нейрон берет начало от больших пирамидных клеток Беца, расположенных в пятом слое коры проекционной двигательной зоны. Аксоны этих клеток идут от коры в глубь полушарий, проходят в составе волокон согопае radiatae белое вещество полушарий и двумя проводниками, т. е. tractus cortico-bulbaris (от нижнего отдела передней центральной извилины) и tractus cortico-spinalis (от верхних двух третей извилины), проходят через внутреннюю капсулу (capsula interna). Внутренняя капсула расположена между большими ганглиями основания; она представляет собой полосу белого вещества, в котором расположены проводники, идущие от коры головного мозга к нижележащим отделам центральной нервной системы, и восходящие проводники, поднимающиеся к коре. Capsula interna делится на переднее бедро (между nucleus caudatus и n. lenticularis), заднее бедро (между nucleus lenticularis и thalamus opticus) и колено (место перегиба, genu capsulae internae, см. рис. 55). Tractus cortico-bulbaris (центральные нейроны для мускулатуры лица, языка, гортани и глотки) проходит в области колена внутренней капсулы; tractus cortico-spinalis, иначе называемый пирамидным пучком, расположен в передних двух третях заднего бедра внутренней капсулы (см. рис. 55, V). В направлении своем книзу оба названных проводника переходят из внутренней капсулы в ножки мозга, занимая средние две трети основания их. Tractus cortico-bulbaris расположен здесь медиально, tractus cortico-spinalis — латерально (см. рис. 53). В мосту (pons Varolii) двигательные проводники располагаются также в основании. Проходящими здесь поперечно и перекрещивающимися волокнами средних ножек мозжечка (tractus ponto-cerebellares) пирамидные пути разделяются на ряд отдельных пучков, сливающихся снова 'в общий проводник в продолговатом мозге. В 'продолговатом мозге пирамидные пучки находятся в основании, обособляясь в два валика, заметные на поверхности мозга — пирамиды (см. рис. 48). На границе продолговатого и спинного мозга пирамидные пучки (tractus cortico-spinales) подвергаются неполному перекресту (decussatio pyramidum). Большая, перекрещенная часть пути переходит в боковой столб спинного мозга и называется основным, или латеральным, пирамидным пучком. Меньшая, неперекрещенная часть проходит в передний столб спинного мозга и носит наименование прямого неперекрещенного пучка (см. рис. 9, 16). Волокна tractus cortico-bulbaris заканчиваются в двигательных ядрах черепномозговых нервов (отсюда другое название проводника — tractus cortico-nuclearis), tractus cortico-spinalis — в передних рогах спинного мозга. Здесь импульсы с центральных двигательных нейронов передаются на периферические. Периферические двигательные нейроны для мускулатуры жевательной, лицевой, языка, гортани и глотки состоят из моторных клеток двигательных ядер черепных нервов с их аксонами, составляющими двигательные волокна корешков и нервов V, VII, IX, Х и XII (тройничного, лицевого, языкоглоточного, блуждающего, подъязычного). Клетки периферических двигательных нейронов для мускулатуры шеи, туловища, конечностей и промежности расположены в передних рогах спинного мозга; аксоны их выходят в составе передних двигательных корешков и составляют двигательные волокна периферических спинномозговых нервов.
Рис. 9. Ход пирамидных путей.
Таким образом, импульсы из коры головного 'мозга проводятся по цепи двух нейронов (центрального и периферического) к мышце. Корковая иннервация мускулатуры — преимущественно перекрестная, так как волокна центральных двигательных нейронов в большинстве своем переходят на противоположную сторону и устанавливают связи с противоположными ядрами черепных нервов и передними рогами спинного мозга. Перекрест волокон tractus cortico-spinalis происходит сразу на границе продолговатого и спинного мозга; волокна же tractus cortico-bulbaris подвергаются перекресту последовательно, по мере подхода их к соответствующим ядрам черепных нервов («надъядерный» перекрест). При поражении центральных двигательных путей расстройства движений, понятно, возникают в мускулатуре противоположной стороны тела, однако не во всей: ряд мышечных групп остается непораженным. Это объясняется наличием так называемой двухсторонней корковой иннервации, существующей для глазодвигательных, жевательных мышц, мышц глотки, гортани, шеи, туловища и промежности. Суть 'в том, что к части двигательных ядер черепных нервов и к некоторым уровням передних рогов спинного мозга волокна центральных двигательных нейронов подходят не только с противоположной стороны, но и со своей, обеспечивая, таким образом, подход импульсов из коры не только противоположного, но и своего полушария. Понятно, что при одностороннем центральном поражении выпадают функции только мышечных групп, односторонне (только из противоположного полушария) иннервируемых. Таковыми оказываются конечности, язык и нижний отдел лицевой мускулатуры, что мы и наблюдаем при так называемой гемиплегии; мускулатура же шеи, туловища, промежности и большая часть мышц, иннервируемых черепными нервами, остается при этом не пораженной. Для осуществления движения необходимо, чтобы двигательный импульс беспрепятственно был проведен из коры головного мозга к мышце. При. перерыве кортико-мускулярного пути проведение импульса невозможно, и соответствующая мускулатура оказывается в состоянии паралича. Неполная утрата движений (уменьшение их силы и объема) называется не параличом, а парезом. По своей распространенности параличи делятся на моноплегии (парализована одна конечность); гемиплегии (паралич одной половины тела), параплегии (поражение двух симметричных конечностей, верхних или нижних); тетраплегии (парализованы все четыре конечности). Параличи или парезы, вызванные поражением отдельных нервных стволов, обозначаются как паралич соответствующего нерва, например, лучевого, локтевого и т.д. Аналогично этому различают параличи сплетений (плечевого, поясничного) или отдельных стволов их. При поражении центральных двигательных нейронов возникает центральный паралич; при поражении периферических нейронов — периферический. Общим для них является лишь самый факт паралича; в остальном симптоматология их резко различается.
Восходящие пути Пучки Голля и Бурдаха (в задних столбах); tractus spino-thalamicus (в боковых столбах); tractus spino-cerebellaris (в боковых столбах).
Нисходящие пути tractus cortico-spinalis lateralis (в боковых столбах); tractus rubro-spinalis (в боковых); tractus cortico-spinalis anterior (в передних столбах); tractus vestibulo-spinalis (в передне-боковых); tractus tecto-spinalis (в передних) и fasciculus longitudinalis posterior (в передних столбах).
Волокна всех нисходящих проводников заканчиваются у клеток передних рогов спинного мозга. Таким образом, периферический двигательный нейрон получает импульсы от всех отделов нервной системы, имеющих отношение к мышечному тонусу и движению. Так, сюда подходят импульсы от центростремительной части рефлекторной дуги, импульсы «произвольного» движения из коры головного мозга по пирамидному пути, от экстрапирамидной двигательной системы по рубро-спинальному и др., от мозжечка по вестибуло-спинальному и тому же рубро-спинальному пучку и т.д. (см. рис. 1 и 16). К периферическому двигательному нейрону подходят, следовательно, как возбуждающие, так и тормозящие импульсы. Здесь, в клетке переднего рога, сосредоточиваются конечные результаты сложной деятельности нервной системы, определяющие и состояние мышечного тонуса и двигательный эффект. Как видно на рис. 16, все перечисленные длинные проводники, связывающие спинной мозг с вышележащими отделами нервной системы, расположены преимущественно на периферии белого вещества. Участки, прилежащие к серому веществу, заняты многочисленными волокнами восходящего или нисходящего направления, то короткими, то более длинными. Это — интерсегментарные или ассоциационные пути, устанавливающие связи между отдельными сегментами спинного мозга. Периферические отделы серого вещества с их ассоциационными клетками и прилежащие отделы белого вещества, где расположены их волокна, составляют так называемый собственный аппарат спинного мозга, связывающий сегменты и обеспечивающий в достаточной мере сложную и разнообразную рефлекторную деятельность спинного мозга. Возникающие при поражении проводникового аппарата спинного мозга расстройства носят, в отличие от сегментарных расстройств, более диффузный характер. При поражении двигательных (пирамидных) путей в состоянии центрального паралича оказываются отделы мускулатуры, иннервируемые от всех нижележащих сегментов. При перерыве чувствительных путей анестезированными оказываются участки, соответствующие также всем расположенным ниже сегментам (так называемые проводниковые расстройства). Поражение заднего столба вызывает утрату суставно-мышечного чувства (и сенситивную атаксию), вибрационной и тактильной чувствительности книзу от уровня поражения на стороне очага. Поражение бокового столба спинного мозга дает книзу от уровня поражения центральный паралич на своей стороне и утрату болевой и температурной чувствительности на противоположной, Половинное поражение поперечника спинного мозга создает картину броун-секаровского паралича, или синдрома. Проводниковые расстройства выражаются в центральном параличе и утрате суставно-мышечного и вибрационного чувства на стороне поражения; на противоположной очагу стороне наблюдается болевая и температурная анестезия[8]. Соответственно пораженным сегментам (серого вещества) на стороне очага могут наблюдаться также сегментарные двигательные (периферические параличи) и чувствительные расстройства[9]. Полное поражение поперечника спинного мозга вызывает параплегию нижних ко
|
||||
|
|
Последнее изменение этой страницы: 2016-04-21; просмотров: 480; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.58.200.78 (0.014 с.) |