
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тема: популяции как простые надорганизменные системыСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
(лабораторное занятие)
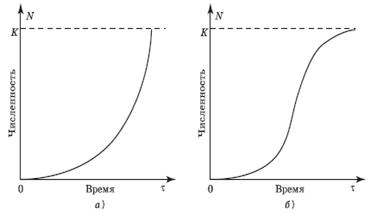
Цель: рассмотреть свойства и динамику популяции как элементарной надорганизменной системы, ознакомиться с методами оценки количественных параметров популяции. Материалы и оборудование: плакаты, калькуляторы, мультимедиа, видеофильм. I. Теоретическая часть Популяция, как отмечалось ранее, представляет собой элементарную надорганизменную систему, объединяющую особей одного вида. Она характеризуется известной устойчивостью во времени и пространстве и относительной обособленностью от других подобных систем. Основу существования популяции составляют закономерные взаимоотношения между особями, в первую очередь связанные с воспроизводством. Свободный обмен наследственным материалом при размножении обеспечивает формирование и поддержание единого популяционного генофонда. Наряду с панмиксúей (случайным, неограниченным скрещиванием),к объединяющим механизмам относятся взаимодействия по типу взаимопомощи, которые особенно ярко проявляются при образовании внутрипопуляционных группировок (семей, стай, стад и др.). Важную роль в регуляции популяционных характеристик играет также внутривидовая конкуренция. Ее напряженность может сглаживаться за счет усложнения этологической (поведенческой) структуры популяции в виде внутривидовой иерархии. Как единица существования и эволюции вида популяция характеризуется рядом параметров, из которых важнейшими можно считать: 1) общую численность, 2) территориальную целостность (при этом особи, образующие популяцию, могут быть распространены по территории равномерно или, что наблюдается чаще, неравномерно – в связи с формированием внутрипопуляционных группировок и неравномерностью распределения ресурсов), 3) плотность (количество особей, приходящееся на единицу площади или объема пространства), 4) внутреннюю упорядоченность. Последняя характеризуется количественными соотношениями особей, различающихся по полу, возрасту, размеру, массе, поведению, генетическим свойствам и т.д. Несмотря на выраженные гомеостатические свойства надорганизменных систем (см. Занятие 1), в практическом плане наиболее важным моментом при изучении популяций разных живых организмов является выявление особенностей их динамики, в первую очередь – динамики численности (плотности). Изменения количественных показателей популяции во времени определяются соотношением рождаемости и смертности особей, а также их перемещениями (эмиграцией и иммиграцией). Динамика численности популяций разных биологических видов может значительно отличаться. Выделяют два основных типа роста популяции: а) экспоненциальный (или J-образный – по форме кривой, построенной в арифметическом масштабе) и б) сигмоидный, который характеризуется S-образной, или логистической, кривой (см. рис. 1). Первый тип описывается простым экспоненциальным уравнением: dN/dt = r×N, где dN/dt – скорость изменения числа особей в единицу времени, r – коэффициент мгновенного прироста популяции (соответствует репродуктивному потенциалу популяции в отсутствие действия лимитирующих факторов среды, то есть при неограниченных ресурсах), N – численность популяции (экз.). Способность к экспоненциальному росту(размножению в геометрической прогрессии) в принципе присуща любому биологическому виду. Однако в случае типичной J-образной кривой плотность популяции возрастает по экспоненте до уровня, при котором возникает нехватка жизненных ресурсов, после чего рост численности достаточно резко прекращается, сменяясь ее падением. Подобная динамика характерна для короткоцикловых, быстро размножающихся видов живых организмов: планктонных водорослей, многих простейших, коловраток, мелких ракообразных (дафний, например), многих насекомых, некоторых рыб и грызунов – которых за развитую способность быстро увеличивать свою численность в благоприятной среде (что соответствует высокому значению показателя r в приведенном выше уравнении) называют r-стратегами. Р. Мак-Артур и Э. Уилсон (1967) даже выделяют особую форму отбора, способствующую выработке подобной стратегии выживания – r-отбор.
Рисунок 1 – Типы кривых роста численности популяции: а – J-образная; б – S-образная; K – поддерживающая емкость среды Другой тип роста популяции – сигмоидный, характеризуется меньшей удельной скоростью, чем экспоненциальный, и описывается логистическим уравнением Ферхюльста-Пирла: dN/dt = r×N×((K-N) /К), в котором присутствует показатель К – предельная численность популяции (верхняя асимптота логистической кривой), соответствующая жизненной емкости среды (запасу жизненных ресурсов). Сигмоидный рост наблюдается в тех случаях, когда скорость увеличения численности популяции определяется, главным образом, действием не абиотических условий, а биотических факторов (факторов, зависящих от плотности), таких, как пресс хищников и паразитов, напряженность меж- и внутривидовой конкуренции и др. С плотностью популяции у живых организмов тесно связаны скорость достижения особями половой зрелости, их плодовитость и ряд других биологических свойств. Типичная К-стратегия, то есть способность поддерживать свою численность на уровне, близком к максимально возможному в данной среде, обнаруживается у долгоживущих, поздно созревающих и медленно размножающихся организмов: крупных хищников, слонов, китообразных, ряда древесных растений и др. На протяжении длительных периодов популяции К-стратегов мало изменяют свою численность (последняя слабо колеблется вблизи значения К), за что их также называют равновесными (видами-регуляторами). Соответственно, сохраняют известное постоянство и другие параметры популяции, в том числе ее генетическая структура, при характеристике которой можно с успехом использовать уравнение Харди-Вайнберга (см. Занятие 6). Напротив, динамика популяций r-стратегов, иначе называемых видами-оппортунистами, характеризуется резкими периодическими или апериодическими колебаниями численности (флуктуациями), амплитуда которых может достигать 3-6 порядков. Для r-стратегов характерна катастрофическая, обычно не зависящая от плотности смертность, проявляющаяся на фоне изменчивой, часто слабой конкуренции, короткой продолжительности жизни и небольшого числа актов воспроизводства. В чистом виде описанные выше типы динамики встречаются сравнительно редко, что позволяет говорить о преобладающей экологической (эволюционной) стратегии, которая для конкретного вида может соответствовать любым переходным вариантам между вышеописанными крайними: r-стратегией, направленной на воспроизведение максимально возможного числа потомков в единицу времени, и К-стратегией, которая ориентирована на максимальное выживание сравнительно немногочисленных потомков. Следует добавить, что динамика численности может отличаться даже у одновидовых популяций, занимающих несходные части видового ареала. Например, выяснено, что многие характеристики r-стратегов обнаруживаются в первую очередь у периферийных популяций, то есть существующих вблизи границ видового ареала (в зоне экологического пессимума). По мере роста использования человеком биологических ресурсов нашей планеты все больше внимания уделяется разработке методов рационального управления природными популяциями. Последнее становится важным условием долговременной и эффективной эксплуатации растительного и животного мира и сохранения живой природы в целом. Только умеренное и избирательное изъятие части особей из популяции промыслового вида (в пределах его репродуктивного потенциала) приводит к увеличению темпов его размножения: за счет роста плодовитости, ускоренного полового созревания, оптимизации внутрипопуляционной структуры и т.п. Следовательно, чтобы поддержать запасы ценных растений и животных, необходимо проведение детальных популяционных исследований эксплуатируемых видов, на результатах которых и должна основываться тактика их рационального использования. Концепция «популяция как единица управления» применима также по отношению к видам, которые с точки зрения человека являются «вредными»: снижают урожай сельскохозяйственных культур, повреждают пищевые запасы, распространяют возбудителей заболеваний или сами вызывают заболевания человека и домашних животных и т.п. Однако в этом случае воздействие на популяции имеет целью не извлечение максимальной устойчивой продукции (как в отношении эксплуатируемых видов), а эффективное снижение численности вредителя (вплоть до его полного уничтожения на выбранной территории в соответствии с принципом девастации, сформулированным академиком К.И.Скрябиным). Осуществление этой задачи сталкивается с рядом проблем. Во-первых, следует помнить, что с точки зрения законов живой природы любой биологический вид является важным элементом ее устойчивого функционирования и потому не может быть «абсолютно вредным». Более того, каждый вид – это уникальный генофонд, и его потеря невосполнима. Во-вторых, подавляющее большинство «вредных» видов относится к r-стратегам: основной ущерб, наносимый хозяйственной деятельности человека такими представителями, связан с неконтролируемыми вспышками их численности. Поэтому наиболее перспективным вариантом «борьбы с вредителями» следует признать тактику «сдерживания роста популяций нежелательных видов», основанную, в частности, на использовании антагонистических взаимоотношений между разными группами организмов в составе единого сообщества (то есть на применении биологических методов «борьбы»). II. Самостоятельная работа Задание 1 Как уже указывалось, к важнейшим характеристикам природных популяций относятся общая численность и плотность. Поскольку прямой учет числа особей в популяциях большинства животных затруднен (вследствие их подвижности), для оценки величины локальной группировки можно использовать индекс Линкольна, который применим в случаях, когда члены популяции распределяются случайным образом, то есть равномерно; при этом численность популяции во времени должна быть достаточно стабильна. Расчет названного индекса предполагает вылов части особей популяции, их мечение и последующий выпуск на волю в месте поимки. Через некоторое время, достаточное для того, чтобы животные могли распределиться случайным образом, снова проводят отлов. Примерный размер популяции определяют по формуле: A=N1×N2/n2, где А – общая численность, экз., N1 – число животных в первом «улове», экз., N2 – число животных во втором «улове», экз., n2 – число помеченных животных во втором улове, экз. Решите задачу. Чтобы оценить численность форели в небольшом озере площадью 125 га, были выловлены 926 форелей, которых путем «выщипа» части спинного плавника пометили и выпустили. Повторно были пойманы 1073 форели, из которых у 129 были обнаружены метки. Определите примерный размер популяции форели в озере и ее среднюю плотность.
Задание 2 Как уже отмечалось, уравнение Харди-Вайнберга [ (p+q)²= p²+2pq+q² (=1), где p – частота доминантного (А), q – рецессивного (а) аллеля; p², 2pq, q² –частоты генотипов АА, Аа и аа соответственно] позволяет оценить генетическую структуру популяции (по одной паре аллельных генов) при условии ее приближенности к идеальной (условно безразмерной, панмиктической популяции, на которую не действуют эволюционные факторы). Решите задачу. У обыкновенной лисицы рыжая окраска шерсти определяется аллелем R, серебристая (черно-бурая) – аллелем r; при этом указанные варианты проявляются фенотипически только у гомозигот, тогда как гетерозиготы (Rr), называемые сиводушками, имеют промежуточную окраску. При исследовании одной из популяций лисицы была учтена 121 особь, из которых оказалось 77 рыжих, 5 чернобурок и 39 сиводушек. Соответствует ли полученное количественное соотношение лисиц с разным цветом шерсти закону Харди-Вайнберга? Контрольные вопросы 1. Что такое популяция? Каковы ее важнейшие характеристики? 2. Охарактеризуйте основные типы динамики численности природных популяций. С какими биологическими свойствами видов они связаны? 3. Каково теоретическое и практическое значение знаний популяционной организации и динамики биологических видов? Занятие 11 Тема: Экосистемы (лабораторное занятие)
Цель: рассмотреть организацию биологического сообщества и биогеоценоза, ознакомиться с процессами, лежащими в основе их существования и динамики. Материалы и оборудование: калькуляторы, плакаты, мультимедиа, видеофильм. I. Теоретическая часть Популяции разных видов живых организмов, совместно существующие на общей территории, образуют биологическое сообщество, или биоценóз (К. Мебиус, 1877). Устойчивость биоценоза в первую очередь основана на закономерных межвидовых взаимодействиях между его членами. Кроме этого, объединяющим моментом является известное сходство требований к величине основных абиотических факторов у составляющих сообщество видов. Главную роль в системе биоценотических связей играют пищевые (трофические) взаимоотношения: хищничество, конкуренция, комменсализм и другие(табл. 6). Хищничество, или биофáгия, является формой взаимодействия двух видов, при которой один из них (хищник) использует другой вид (жертву) в качестве источника питания. Биофагию делят на растительноядность и животноядность (собственно хищничество). Конкурентные отношения между организмами возникают при использовании одинаковых ресурсов. Результатом конкуренции является снижение доступности ресурса (пищи, в частности) для каждого из взаимодействующих видов. Стоит заметить, что напряженная межвидовая конкуренция в конечном итоге приводит к вытеснению одного из конкурирующих видов другим; данная закономерность известна в биологии как принцип конкурентного исключения, или закон Г.Ф.Гаузе. Комменсализм, или нахлебничество, предполагает извлечение односторонней выгоды (для комменсала). Так, остатки несъеденной хищником жертвы часто достаются различным «падальщикам». Таблица 6 – Основные типы взаимоотношений между популяциями двух видов в составе биологического сообщества *
*Примечание: (+) означает извлечение видом выгоды из существующих связей; (-) означает ущерб (вред), испытываемый популяцией в ходе взаимодействия; (0) – отсутствие значимых последствий от взаимодействия (отсутствие значимого взаимодействия). Линейная последовательность живых организмов, в которой каждое предыдущее звено представляет источник питания для следующего, образует трофическую цепь. Выделяют два типа пищевых цепей: пастбищные (цепи выедания) и детритные (цепи разложения). Первые начинаются с автотрофных организмов, в качестве которых обычно выступают растения, создающие органическое вещество из неорганического в ходе фотосинтеза. Их называют продуцентами. Второе и последующие звенья пастбищной пищевой цепи представлены гетеротрофными организмами (консументами), потребляющими готовый органический материал, созданный на предыдущем уровне, и преобразующими его в вещества собственного тела. Роль консументов в трофической структуре сообщества, как правило, выполняют животные. В зависимости от характера питания и положения в пищевой цепи их делят на первичных (растительноядных), вторичных и третичных (плотоядных) консументов. Замыкают пищевую цепь редуценты – разрушители органического вещества (минерализаторы), каковые чаще всего обнаруживаются среди сапрофитных бактерий и грибов. Следует заметить, что в разрушении мертвой органики существенную роль также играют животные- детритофаги, некрофаги и копрофаги. Детритные цепи начинаются и заканчиваются сапротрофами (детритофагами). Наболее характерны они для лесных сообществ и донных (бентических) биоценозов глубоководных озер и океанов. Кроме трофических, важной группой биоценотических связей между популяциями разных видов являются топические, связанные с совместным использованием пространства. Под ними понимают любое изменение (физическое или химическое) среды обитания одного вида в результате жизнедеятельности другого. Примером подобных связей являются квартиранство (норовый и гнездовой комменсализм), разные варианты паразитизма, в первую очередь внутреннего, или эндопаразитизма (при этом не стоит забывать, что паразит использует хозяина также в качестве источника питания), и аменсализм. О последней форме связей говорят в тех случаях, когда какой-либо вид испытывает негативное влияние другого, который, в свою очередь, не ощущает сколько-нибудь значимого воздействия со стороны первого. Так, высокие деревья в лесу затеняют низкорослые кустарники и травы. Кроме названных выше, в структуре связей биологического сообщества выделяют также форические связи, имеющие форму участия одних организмов в распространении других, и фабрические взаимодействия. Последние связаны с использованием многими животными частей тела (шерсти, перьев, листьев, стеблей и др.), продуктов жизнедеятельности или мертвых остатков других видов для целей «строительства» гнезд, нор, домиков и т.п. Как уже отмечалось в предыдущей теме, антагонистические взаимоотношения, в частности хищничество, паразитизм и конкуренция, участвуют в регуляции популяционных показателей участников сообщества. Одной из важнейших характеристик биоценоза является его видовая структура. При ее определении значимым является не только общее число видов (видовое богатство), но и их количественное соотношение (выравненность). Используя таксономические подходы, биологическое сообщество делят на фитоценоз (растительное сообщество), зооценоз и микробоценоз. Отдельные доминирующие члены биоценоза (в первую очередь это растительные формы) могут выполнять важную средообразующую роль (оказывают сильное топическое воздействие на другие виды сообщества). Их называют видами- эдификаторами. Помимо видовой структуры, биоценоз как сложная система характеризуется пространственной структурой в форме ярусности (вертикальной слоистости) и мозаичности (горизонтальной неоднородности), трофической структурой (в виде сложной суммы пищевых цепей – пищевой сети), комплексом взаимовыгодных межвидовых взаимодействий и сформированной на его основе коадаптивной структурой и т.д. Немецкий эколог В. Тишлер (1971) в качестве основных особенностей биоценоза как саморегулирующейся системы выделил следующие: 1) части сообщества взаимозаменяемы; один вид может быть заменен другим со сходными экологическими свойствами без какого-либо ущерба для устойчивости самой системы; 2) сообщество основано на количественной регуляции одних видов другими; 3) сообщество поддерживается главным образом за счет уравновешивания противоположно направленных сил – антагонистических взаимоотношений между его членами. Физическое пространство с определенным набором абиотических условий, в пределах которого существует биологическое сообщество, называется биотóпом. Процессы извлечения вещества и энергии из внешней среды живыми организмами, их последующее превращение в пищевых цепях биоценоза и возвращение в абиотическую среду в виде продуктов метаболизма объединяет биоценоз и биотоп в ещё более сложную систему – биогеоценоз (В.Н. Сукачев, 1940). Совокупность всех биогеоценотических связей какого-либо вида организмов определяется понятием экологическая ниша (Ч. Элтон, 1927; Дж. Хатчинсон, 1957). Последняя включает также пространственное размещение вида в биологическом сообществе, однако нишу следует отличать от «местообитания» (стáции), под которым понимают участок пространства, занятый группой особей некоего вида и обладающий необходимыми для ее существования условиями. Местообитание, следовательно, является компонентом экологической ниши вида (можно еще раз напомнить, что у многих животных со сложным жизненным циклом наблюдается онтогенетическая смена стаций). Таким образом, экологическая ниша характеризует положение вида в экосистеме и весь комплекс его требований к окружающей среде. Все биогеоценозы нашей планеты объединяются в крупнейшую (глобальную) живую систему Земли – биосферу. Основу ее существования, как и основу функционирования отдельного биогеоценоза, составляет движимый энергией Солнца биотический круговорот, который можно представить как совокупность биогеохимических циклов химических элементов, входящих в состав живой клетки. Биогеохимические циклы делят на газовые (с резервным фондом химических элементов, расположенным в атмосфере или гидросфере) и осадочные с резервным фондом преимущественно в земной коре (см. Приложение Г). Первые отличаются от вторых большей скоростью и стабильностью. Для газовых циклов характерна также бóльшая степень замкнутости, хотя круговорот любого биогенного элемента никогда не бывает полностью обратимым (максимум до 98 %). Поглощенная продуцентами солнечная радиация (ФАР) в ходе метаболизма трансформируется в энергию химических связей органических соединений. Об эффективности усвоения энергии можно судить по скорости создания органического вещества, то есть по величине продукции. При переходе с одного трофического уровня на каждый последующий количество энергии, первоначально ассимилированной автотрофами в виде первичной продукции (ассимиляционная эффективность фотосинтеза составляет в среднем около 1 %), закономерно снижается в 5-10 раз. Это обусловлено тем, что любой организм бóльшую часть поглощённой извне энергии тратит на поддержание основного обмена, превращая её в конечном счёте в тепло. Консументы, кроме того, значительный объем химически связанной энергии теряют с экскрементами. По этой причине число звеньев в отдельной пищевой цепи всегда ограничено (обычно не превышает 4-6). Чистая продукция экосистемы, определяемая как суммарный прирост биомассы живых организмов в единицу времени, обратно пропорциональна ее гомеостатическим свойствам. Эволюционные (в первую очередь эндоэкогенетические, то есть вызванные внутренними свойствами самой системы) изменения биоценоза (биогеоценоза), называемые сукцессиями, направлены в сторону усложнения структуры и увеличения общей биомассы. Это сопряжено с ростом затрат энергии на поддержание устойчивого круговорота вещества и, следовательно, со снижением чистой продукции. Различают первичные сукцессии, протекающие на участках, не затронутых почвообразованием (скалах, речных наносах, сыпучих песках и т.п.), и вторичные, которые наблюдаются на месте ранее существовавших, но подвергнувшихся разрушению биоценозов. В результате сукцессионной смены возникает зрелое, или климаксное, сообщество, находящееся в относительном равновесии со средой. Искусственные экосистемы, в частности экосистемы сельскохозяйственного назначения – агробиоценозы (поля, сады, искусственные пастбища, животноводческие фермы, рыбоводные пруды и т.д.) характеризуются упрощенной видовой и трофической структурой, что обеспечивает высокий выход продукции (урожайность), но слабые способности к саморегуляции. Повышение устойчивости агроценозов без потери количества и качества производимой ими биопродукции – насущная задача сегодняшнего дня. Достижению этой цели, в частности, служит внедрение в ряде отраслей сельского хозяйства (например, в растениеводстве, рыбоводстве и др.) поликультуры – приема одновременного культивирования в рамках единого агроценоза нескольких специально подобранных видов (положительно воздействующих друг на друга) и так называемых интегрированных технологий. II.Самостоятельная работа Задание 1 Одной из важнейших частей синэкологических (экосистемных) исследований является выявление видовой структуры биоценоза, или, иначе, видового разнообразия (альфа-разнообразия). Как уже было отмечено, при его оценке обычно учитываются два показателя: видовое богатство – общее число видов в сообществе (количество видов, отнесённое к определённой площади, называется видовой плотностью), и выравненность – равномерность распределения видов по их обилию (доле) в сообществе. Выравненность максимальна, если все виды в сообществе одинаково обильны, и минимальна, когда численность одного вида (доминирующего) превышает численность всех остальных. Высокую выравненность принято считать эквивалентной высокому биоразнообразию. Исследователь обычно не располагает полным списком видов сообщества, а имеет дело с выборкой. Следует помнить, что при увеличении объёма выборки число видов, в ней представленных, как правило, растёт. Одним из простейших показателей биоразнообразия является индекс видового богатства Маргалефа (Лебедева и др., 2004): D = (S – 1)/ln N, где S – число выявленных видов, N – общее число особей всех видов (экз.). Решите задачу. Рассчитайте величины индекса Маргалефа на основе результатов исследования разнообразия птиц в театральном парке г. Ростова-на-Дону, где выборка объемом в 149 особей включала 17 видов птиц, и в ЦПКиО г. Киева, где были учтены 254 особи 19 видов птиц. В каком случае видовое разнообразие оказалось выше? Задание 2 В синэкологических исследованиях чаще используются индексы, учитывающие одновременно и видовое богатство, и относительное обилие (доли) видов, которые иначе называют индексами неоднородности. К числу индексов неоднородности, полученным на основе теории информации (информационно-статистическим), относится индекс разнообразия Шеннона (1949), рассчитываемый по формуле: H′= -Σ pi × ln pi, где pi – доля особей определённого (i-го) вида, которая вычисляется как ni /N, при этом ni – число особей i-го вида в выборке (экз.), N – общее количество особей всех видов (экз.). Необходимо заметить: использование индекса Шеннона предполагает, что особи в выборку попадают случайно из «неопределённо большой» генеральной совокупности, причём в выборке представлены все виды сообщества. Решите задачу. Рассчитайте индексы Шеннона по результатам изучения ихтиоценозов двух участков Воткинского водохранилища на р. Каме в 1981 г. (см. табл. 7); сравните видовое разнообразие рыб в указанных участках. Таблица 7 – Численность рыб разных видов в экспериментальных сетных уловах в Воткинском водохранилище в 1981 г. (экз).
Контрольные вопросы 1. Назовите основные типы взаимодействий между видами организмов в составе биологического сообщества. Приведите примеры. 2. Какие части включает биогеоценоз? Что их связывает? 3. Охарактеризуйте структурные показатели (типы структур) биоценоза (биогеоценоза). 4. Опишите типы и закономерности сукцессионных изменений биогеоценоза. 5. Чем различаются биогеоценоз и агроценоз? Какие возможности повышения устойчивости агроэкосистем вам известны? Приложения
Приложение А
Приложение Б
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-18; просмотров: 895; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.119 (0.019 с.) |

 Основные методы, используемые в биологии
Основные методы, используемые в биологии





