

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
О физиологических механизмах поведения простейших
Об условных рефлексах простейших. Рассмотрение оригинальных экспериментальных исследований, посвященных выработке у простейших условных рефлексов, убеждает в чрезвычайно разноречивой трактовке результатов. Обзору таких работ посвящен ряд публикаций, отражающих взгляды биологов зоологического и физиологического профилей (Коган, 1963; Воронин и др., 1967; Серавин, 1969, 1978; Hamilton. 1975; Тушмалова, 1980). Исследователи, иллюстрирующие возможность выработки, условных рефлексов у простейших, чаще всего останавливаются на опытах Н. Н. Тимофеева (1958) и серии работ Гельбера (Gelber, 1956, 1958). Рассмотрим детально эти эксперименты, чтобы показать, что их результаты могут иметь иное объяснение, если учесть некоторые особенности ресничных инфузорий. Н. Н. Тимофеев проводил опыты на инфузориях в камере размером 10x4 мм, разделенной пополам, в каждой из половин были вмонтированы платиновые электроды. Условным раздражителем служил свет, безусловным — электрический ток (50 Гц 1 — 2 В). Выработка условного рефлекса заключалась в том, что инфузории "учились" только в ответ на изолированное действие света не заплывать в ту часть камеры, где давался удар электрического тока. Объясняя результаты опытов автор акцентирует внимание на двигательных реакциях инфузорий, считая, что выработка условных реакций определяется "количеством, а возможно, и качеством самой двигательной активности", не раскрывая этого понятия, однако опыты Н. Н. Тимофеева можно объяснить исключая возможность формирования временных связей. Известно, что многие инфузории в ответ на различные вредные воздействий внешней среды выстреливают трихоцисты 1. Эту реакцию можно классифицировать как врожденную, безусловно оборонительную — сигнал "опасности" для других особей. В опытах Н. Н. Тимофеева таким "оборонительным" раздражителем был электрический ток. Формирование "условного рефлекса" во времени (постепенность выработки) на самом деле может быть следствием постепенного диффундирования относительно небольшого количества трихоцист по объёму камеры. Экспериментально показано, что выстреливание трихоцист происходит в ответ на непосредственное действие различных кислот, щелочей, механических воздействий электрического тока (Prosser, Brown, 1967).
Таким образом, результаты опытов Н. Н. Тимофеева не могут служить доказательством способности простейших вырабатывать условные рефлексы.
Часто как доказательство выработки у простейших условных рефлексов приводят серию работ Гельбера по выработке у инфузорий Paramecium aurelia пищевых условных рефлексов. В связи с принципиальной важностью физиологической оценки этих работ в плане обсуждаемой проблемы, остановимся подробно на рассмотрении результатов этих работ. Опыты проводились следующим образом. Стерильная платиновая проволока длиной 7,6 см с диаметром кончика 0,5 мм располагалась в середине стеклянной экспериментальной лунки под углом 85° к горизонтали. Поднимание и опускание проволоки осуществлялось с помощью специального рубильника. В контрольных экспериментах было показано, что погружение проволоки в нативную культуру 2 на 3 мин приводит к скоплению на ней 1 — 2 инфузорий. При обучении проволоку опускали в лунку с голодными инфузориями на 15 с. с интервалами в 25 с (время отсчитывали фотографическим хронометром).
Каждое третье опускание проволоки подкреплялось пищей (опускалась проволока с нанесенными бактериями). В контрольных экспериментах не было обнаружено изменений в количестве животных, что исключило возможное объяснение увеличения числа животных у проволоки за счет простого изменения двигательной активности инфузорий.
Однако в этой работе не обнаружено воздействия обучения с подкреплением, что, по мнению автора, может быть объяснено слабой или замедленной реакцией животных на эффект опускания проволоки. Хотя такое объяснение автора и не лишено основания, более вероятной причиной нам кажется нарушение процедуры выработки условных рефлексов: слишком большие интервалы между сочетаниями (оптимальные условия частоты вообще специально не изучались) и непериодичность подкрепления (лишь третье предъявление иглы сочеталось с пищей). В дальнейшем Гельбером было показано, что выработанная реакция инфузорий — прилипание "дрессированных" животных к стерильной проволоке — сохраняется до 3 ч. Был продемонстрирован эффект, аналогичный угасанию, у трех групп культур P. aurelia, обученных по описанной методике, через 2 ч проверялось сохранение обучения. Группы перед началом проверки получили разное количество испытаний без подкрепления: группа 1 — 10 испытаний, группа 2 — 5, группа 3 — не получала совсем. Оказалось, что через 2 ч после дачи проб группа 3 показала количество ответов, превышающее остальные группы. Группы без обучения (контрольные) дали "нулевые" ответы. Эти опыты, без сомнения, подтверждают пластичность поведения инфузорий. Однако они, как и предыдущие эксперименты автора, не дают (в силу особенностей используемой методики) ответа на природу физиологических механизмов, лежащих в основе наблюдаемого явления. Отмеченное угасание на самом деле могло развиваться по механизму привыкания к отсутствию пищевого раздражителя, что исключает условнорефлекторную природу явления.
В этой связи заслуживают внимания опыты Катца и Детерлайна (Katz, Deterline, 1958), которые использовали методику Гельбера с дополнительными контрольными экспериментами и показали, что после перемешивания жидкости в лунке с бактериями "дрессированные" бактерии не оседают на поверхности стерильной проволоки на дне лунки. В другом опыте авторы на дне лунки с голодными парамециями вносили с помощью платиновой проволоки небольшое количество бактерий. Повторив методику выработки условных рефлексов у инфузорий P. aurelia, они использовали только стерильную платиновую проволоку. Результаты достоверно опровергали опыты Гельбера: были получены такие же данные, как и в случае с проволокой, покрытой предварительно бактериями (инфузории без "обучения" собирались на поверхности стерильной проволоки).
Таким образом, результаты опытов Гельбера, на наш взгляд, могут свидетельствовать лишь о способности инфузорий четко реагировать на пищевое раздражение или, возможно, на изменение среды, вызванное присутствием бактерий.
Привыкание простейших. У организмов, лишенных нервной системы, могут быть более простые (неусловнорефлекторные) формы приспособительного поведения. Примером элементарного механизма накопления индивидуального опыта служит привыкание (Thorp, 1964). Под привыканием понимают прекращение реакции на постоянно действующий биологический раздражитель. Л. Г. Воронин (1968, 1969, 1972) относит привыкание простейших к несигнальной форме индивидуального приспособления. К одной из первых работ по выработке привыкания у простейших принадлежат опыты Данича (Danisch, 1921) на сувойках (Vortioella nebulivera). Автор исследовал привыкание к механическому раздражению у инфузорий. Критерием привыкания служило прекращение сокращения стебелька сувойки в ответ на раздражение. О динамике привыкания судили по количеству сокращений, необходимых для отсутствия сокращения. В работе получена зависимость скорости выработки привыкания от силы механического раздражения — привыкание вырабатывалось тем быстрее, чем меньше была сила используемого механического раздражителя.
Позднее закономерности привыкания у простейших изучал Кинастовский. Им было подробно изучено влияние механического раздражения на сокращение инфузорий (Kinastowski, 1963). Автор в качестве механических раздражителей использовал свободно капающую каплю и специальный прибор, вызывающий вибрацию экспериментального сосуда. Оказалось, что оптимальными условиями для развития привыкания служит сила раздражителя от 400 до 1 600 эрг, частотой 10 и 15 раз/мин. Раздражение с большей силой (20 000 эрг) не приводит к уменьшению числа сокращений, а может вызывать судорожное состояние и летальный исход. Действие раздражителя оптимальной силы с частотой 1 раз в 1 мин не снижало числа сокращений во времени. Полное прекращение реакций в ответ на механическое раздражение в оптимальных условиях опыта наступало через 13 — 20 мин.
В опытах Кинастовского животные с выработанным привыканием отвечали на сигнал другой модальности и на сигнал этой же модальности, но большей силы, что отличало полученный феномен от утомления. В аспекте рассматриваемых работ представляют интерес опыты А. Б. Когана (1963, 1964) на сувойках. Была показана способность этих инфузорий сокращаться в ответ на механическое раздражение — падение на предметный столик маленького свинцового шарика с высоты 3 см. Оказалось, что если к телу сувойки подвести стеклянную палочку так, чтобы выпрямление стебелька осуществлялось только на 3/4 его длины, то инфузории могут изменять свое поведение (замедление выпрямления стебелька, сокращение длины при выпрямлении) и, что особенно важно, сохранять эти изменения некоторое время после прекращения раздражения — удаления палочки. Автор опытов называет такую форму поведения своеобразной приспособительной реакцией. Отсутствие условного сигнала делает невозможным предположение об условнорефлекторном механизме формирования следовых реакций в этом случае. Можно предположить, что у сувоек вырабатывалось привыкание к модифицированному сокращению.
Влияние различных факторов на выработку привыкания у инфузорий спиростом исследовал Аппельвайт (Applewhite, Morowitz, 1967; Applewhite, 1968; Applewhite, Stuart, 1969). В опытах Аппельвайта изучалось привыкание к вибрационному раздражению на популяциях из 60 — 100 инфузорий. Вибрационный раздражитель подавался каждые 4 с (падение груза с силой в 70 условных единиц). Критерием привыкания служило отсутствие сокращений на три раздражения подряд (использовалась фоторегисграция). Сохранение выработанного привыкания проверялось через 15 и 30 с после окончания опытов. В процессе опытов выявилась зависимость привыкания от температуры: при проверке сохранения привыкания через 15, 30, 60 и 90 с при температурах +15, +25 и +37° С оказалось, что более быстрое забывание (больший процент сократившихся инфузорий) наблюдается при температуре 37 С. Известно, что среда культивирования простейших может оказывать влияние на некоторые показатели функционального состояния животных (биение ресничек, изменение вязкости протоплазмы и т. п.). Поэтому особый интерес представляют эксперименты Аппельвайта и сотрудников (Applewhite et al., 1969) по изучению влияния на привыкание ионов металлов. Оказалось, что предварительное 15-минутное содержание инфузорий в растворе хлоридов калия и натрия (в концентрации к 0,5 до 0,005 моль/л), а также кальция, магния и марганца (в концентрации от 0,25 до 0,002 моль/л) не повлияло на выработку и сохранение привыкания, а воздействие ионов магния оказалось ярко выраженным. В этих опытах о динамике привыкания судили по первой, средней и последней парам раздражений, сохранение проверялось через 30 и 120 с после воздействия последнего раздражения. Двигательная активность "магниевых" инфузорий при этом не изменялась. Одним из предполагаемых механизмов влияния магния на привыкание авторы считают активирование эти ионом некоторых энзимов 3.
Интересные данные получены Аппельвайтом и соавторами (Applewhite et al., 1969) при изучении клеточной локализации феномена привыкания. После выработки привыкания животных разрезали пополам и вновь отдельно обучали переднюю и задние части. Оказалось, что для принятого критерия выработки (отсутствие реакции до трех нулей подряд) понадобилось примерно одинаковое число механических стимулов. Анализ полученных фактов позволяет согласиться с мнением авторов работы о том, что привыкание не является следствием утомления, повреждения или местной адаптации.
Изучение роли макронуклеуса в процессе привыкания продемонстрировало отсутствие различий в динамике выработки этой реакции у половинок инфузорий, содержащих ядро и лишенных его, что позволило автору высказать предположение о локализации памяти клетки в цитоплазме. Оценивая серию работ Аппельвайта и сотрудников по изучению привыкания у ресничных инфузорий, необходимо отметить, что они вырабатывали нестойкие реакции (отсутствие реакции на три последовательных стимула), которые при оценке динамики выработки могут быть определены как первые признаки формирования привыкания, но не как прочно выработанная реакция.
Основные закономерности выработки привыкания у Stentor coeruleus были изучены Вудом (Wood, 1970, 1972). У животных вырабатывалось привыкание к механическому раздражению. При постоянной частоте раздражения увеличение силы сигнала замедляло скорость выработки привыкания: в опытах с постоянной силой тока привыкание вырабатывалось тем медленнее, чем реже давался сигнал; при очень сильных раздражениях привыкание не вырабатывалось совсем. Следы привыкания в виде снижения частоты ответов на механические стимулы сохранялись в течение 3 ч.
Несомненным достоинством опытов Вуда служит использование им теста на "растормаживание" как одного из существенных показателей наличия или отсутствия обучения в такой элементарной форме, каким является привыкание. Оказалось, что привыкание, выработанное на механический стимул с силой в 0,04 усл. ед., не растормаживается после однократного раздражения большей силы (0,12 усл. ед.) или после подпорогового раздражения электрическим током. По мнению автора, неспособность к растормаживанию служит единственным отличием свойств привыкания одноклеточных организмов от высших животных.
Оригинальную форму обучения у стентора наблюдали Беннет и Френсис (Bennet, Francis, 1972). Критерием обучения служило сокращение времени передвижения стентора в капиллярной трубке, соединенной с относительно большим резервуаром, наполненным водой. При повторении проб с частотой один раз в 60 с время сокращалось от 58 с в первой пробе до 31 с в третьей. Реакция сохранялась примерно на одном уровне в течение 30 мин. Было показано также, что обучение происходит лишь при строгом соблюдении определенных условий опыта — вертикальном расположении капиллярных трубок с внутренним диаметром не более 0,5 мм. В трубках с диаметром 1 и 2 мм обучения не происходило. Авторы приходят к выводу о том, что изученный вид обучения есть не что иное, как результат привыкания стентора к механическому стимулу. Знакомство с экспериментами Беннета и Френсиса позволяет нам подобное объяснение считать справедливым.
Итак, приведенный материал показывает, что одноклеточные организмы обладают свойством изменять поведение под влиянием различных раздражителей. Эти изменения сохраняются в течение 30 — 60 мин, а иногда до 3 ч. Сохранение следов от раздражений свидетельствует о способности простейших к накоплению индивидуального опыта.
Наиболее детально вопрос о характерных особенностях функциональных механизмов, определяющих поведение простейших, был исследован в серии работ Н. А. Тушмаловой с сотрудниками (Тушмалова, 1968, 1974, 1977; Тушмалова, Доронин, 1971; Доронин, 1974; Тушмалова, Зазулина, 1977; Доронин, Тушмалова, 1978; Тушмалова, Кузьмичева, 1979). Приступая к изучению привыкания у донервных животных на примере привыкания Spirostomum ambigaum к вибрационному раздражению, авторы поставили своей целью прежде всего ответить на вопрос: в какой степени привыкание простейших соответствует понятию приобретенное поведение, выработанному в опытах на позвоночных животных и высших беспозвоночных. Были изучены такие свойства привыкания, как тренированность, растормаживание, информационная значимость интервалов между раздражениями, зависимость выработки этого феномена от исходного функционального состояния и возраста животных. По динамике реакции привыкания всех животных можно отнести к двум группам. В первой группе привыкание было относительным и заключалось лишь в уменьшении числа сокращений в единицу времени. Во второй группе животных реакция была абсолютной — в ответ на раздражение сокращения отсутствовали. Первое проявление привыкания отмечено после 1 — 10 мин; отсутствие сокращений до 10 нулей подряд — после 13 — 47 мин. Как показал анализ экспериментальных данных, оптимальными следует считать сигналы, действующие через промежутки времени, сравнимые с рефрактерными периодами 4 (около 5 с), а по силе — с порогом сокращения инфузорий. Быстрее всего привыкание развивается при действии сигнала через 7 с. При увеличении интервала между раздражениями увеличивается время, необходимое для достижения 100 %-ного угашения реакции. Однако реакция вырабатывалась и при сигнале, действующем значительно реже — через 1 мин: в этих случаях феномен развивался значительно медленнее.
Привыкание вырабатывалось и при раздражении 1—9 раз в минуту. Однако для достижения критерия 10 нулей требовалось значительно больше времени (до 50 мин). Наиболее четко значение временного интервала в динамике выработки привыкания было продемонстрировано в опытах с применением раздражителей в аритмическом режиме, по случайному закону, отмечалось замедление в выработке привыкания при аритмическом режиме раздражения. Как и у более высокоорганизованных животных, у инфузорий были выявлены индивидуальные отличия в динамике привыкания (рис. 1). У животных с выработанным привыканием в 100 % сохранялась реакция на сигнал другой модальности: и более сильный сигнал этой же модальности, что отличает изучаемый феномен от утомления. Условия, при которых наблюдалось утомление инфузорий, резко отличались от условий выработки привыкания. Нереагирование на сигнал любой силы и частоты — утомление — развивалось в результате длительной стимуляции инфузорий до 3 — 4 ч с частотой раздражения 1 раз в 1 с и силой, превышающей пороговую в 4—5 раз.
При изучении ультраструктурных изменений инфузорий в процессе привыкания также обнаружены разнонаправленные сдвиги при привыкании и утомлении в ядрышке макронуклеуса. Так, привыкание сопровождалось активацией ядрышка — увеличением синтеза рибосомной РНК. При утомлении наблюдалось резкое снижение синтеза рибосомной РНК.
Анализ результатов показал, что привыкание к вибрационному раздражителю может быть отнесено к категории явлений, характеризующих следовые процессы, т.е. к памяти. Неясным оставался вопрос о том, в какой степени следовые явления одноклеточных обладают свойствами, общими с другими, более высокоорганизованными животными. Одним из критериев приобретенности (выработанности) реакции служит способность к растормаживанию 5. Использование приема растормаживания привыкания показало, что эта реакция никогда не восстанавливается при действии экстренных раздражителей более слабых, чем агент, на который было выработано привыкание. При действии же раздражителей большей силы, чем индифферентный, растормаживание носило случайный характер. Воздействия стимулами другой модальности (электрический ток, культуральная среда, насыщенная кислородом или углекислотой) не дали положительных результатов. Эффект растормаживания привыкания был отмечен лишь при действии очень сильного раздражителя — пищевого, т. е. при внесении в камеру с инфузориями капли бактериальной суспензии. Для выяснения вопроса, какой же из факторов (рН среды или непосредственно бактерии) является действующим, в контрольных экспериментах в камеру вносили каплю прокипяченной суспензии либо фильтрат суспензии бактерий, либо чистый подщелоченный культуральный раствор. Статистически достоверное растормаживание наблюдалось только при использовании капли культуральной среды повышенной щелочности (рН > 8).
Помимо растормаживания, важным критерием способности животных формировать следовые реакции: является ускорение обучения при неоднократном повторении процедуры обучения — тренированность. У популяции инфузорий привыкание вырабатывалось в течение 20 мин, затем инфузории отсаживались в часовое стекло с культуральной жидкостью. Через 60 мин клетки вновь подвергались воздействию вибрационного раздражители Процедура повторялась 5 — 6 раз. Анализ полученных данных показал, что при первом применении раздражителей (первый опыт) уровень ответов снижается до 35—40 %. По мере повторения опытов достоверно уменьшается вероятность сокращения только на первые раздражители. При 5-й и 6-й повторностях инфузории реагируют на механический стимул с вероятностью, которая не отличается (Р > = 0,9) от уровня ответов в конце первого опыта. Таким образом, процесс тренированности привыкания у инфузорий выражается только в понижении вероятности сокращения на первые стимулы по мере применения серий раздражителей (при интервале между опытами 60 мин).
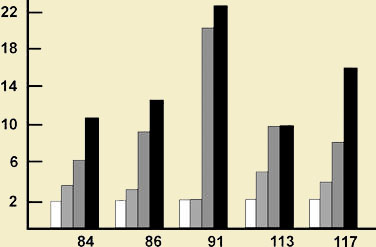
Рис. 1. Динамика выработки привыкания у отдельных инфузорий (по: Тушмалова, 1972): по оси ординат — время, мин; по оси абсцисс — номера инфузорий; светлые столбцы диаграмм — время, необходимое для достижения 1 нуля, с сетчатой штриховкой — 3, с косой штриховкой — 5, черные — 10 нулей
Не отмечено прямой зависимости динамики привыкания от уровня спонтанной двигательной активности и пищевой возбудимости: скорость выработки не зависела от числа пищеварительных вакуолей. Однако при длительном голодании привыкание не вырабатывалось. Обнаружена зависимость привыкания от возраста инфузорий и состояния их ядерного аппарата. Оптимальным для формирования привыкания оказался возраст 45 — 55 ч после деления (примерно 50 % продолжительности жизни). В возрасте 105 ч, когда макронуклеус инфузорий находится в состоянии конденсации (физиологическая реорганизация, по И. Б. Райкову, 1967), привыкание не вырабатывалось.
Говоря о привыкании простейших, важно подчеркнуть, что этот феномен проявляется не только у ресничных инфузорий, но и у более простых одноклеточных — Amoeba proteus. Н. А. Тушмалова и И. Л. Кузьмичева (1978, 1979) изучали закономерности формирования привыкания амеб на периодическое предъявление света 800 — 2 000 лк с интервалом в 1 — 2 мин. Критерием выработки привыкания служило прекращение течения цитоплазмы в ответ на 10 предъявлений света подряд. Как и у ресничных инфузорий, у амеб было выявлено абсолютное привыкание (52,5%) и относительное — уменьшение числа ответных реакций (47,5%). Оказалось, что по зависимости скорости выработки привыкания от силы и частоты раздражения этот феномен у амеб сопоставим с таковым у спиростом.
Таким образом, проведенные опыты показали, что приобретенное поведение простейших обладает свойствами, общими с таковыми у животных, имеющих нервную систему, и одновременно несет черты более примитивной организации.
Поведение кишечнополостных
Об условнорефлекторной деятельности кишечнополостных. Вопрос о способности кишечнополостных формировать условные рефлексы в настоящее время остается открытым. Результаты немногочисленных экспериментальных попыток выработки условных реакций у различных видов этого типа до сих пор не дали положительных результатов. Одна из первых работ была предпринята А. А. Зубковым и Г. Г. Поликарповым (1951). Они проводили опыты в часовом стекле и обнаружили, что движения гидры, прикрепленной близко к поверхности воды, совершаются в направлении от поверхности воды, хотя на глубине в условиях равномерного освещения наблюдаются движения во всех направлениях; через 60 мин после начала опыта движения в сторону поверхности воды прекращаются. При вторичном повышении уровня воды "выработанная" реакция сохраняется до 3—4 ч, а затем угасает. На основании таких свойств реакции, как угашение при неподкреплении, торможение при действии экстрараздражителей (сотрясение и изменение освещенности), авторы определили наблюдаемую модификацию поведения гидр как выработку условного рефлекса, однако результаты проведенных опытов могут иметь и иные объяснения. Так, Л. М. Чайлахян (1957) объясняет "приобретенность" такой реакции гидр лишь изменением состояния мышечных элементов. Более детальный анализ условнорефлекторной деятельности кишечнополостных дан в работе Л. М. Чайлахяна (1957) на примере опытов с пресноводным полипом. Сочетание света (условный сигнал длительностью от 5 с до 1 мин) с электрическим током — безусловный раздражитель при интервалах от 2 до 3 мин привело к проявлению реакции сокращения животных только на изолированное действие света. При увеличении интервала между условными и безусловными раздражителями до 5—6 мин "условная" реакция отсутствовала. Выявленные закономерности позволили автору заключить, что реакция полипа на свет не условный, а суммационный рефлекс — результат повышения возбудимости животного вследствие суммации следов от предыдущих раздражений электрическим током, под влиянием чего допороговая интенсивность света становилась пороговой.
Нами была проведена серия экспериментов по изучению способности к формированию условнорефлекторной памяти у пресноводной гидры (Hydra attenuate). В первых двух сериях опытов (Тушмалова, Симирский, 1977) были исследованы реакции гидр на действие сочетанных раздражителей при попытке выработать пищевые и электрооборонительные условные рефлексы. Условным сигналом служил свет лампы освещенностью в 1000 лк с изолированным действием в течение 30—60 с, на протяжении 30 с совпадающий с безусловным пищевым подкреплением (циклопы). Интервалы между сочетаниями определялись временем заглатывания циклопа и составляли 6 — 10 мин. Критерием положительной условной реакции было выбрано открывание гипостома в ответ на изолированное действие света. Контролем служили животные, которые ежедневно получали дозированную пищу в отсутствии света, и животные, получающие пишу при непрерывном освещении красным светом. Изменение реакции гидр от сочетания к сочетанию выражалось лишь в том, что свет неспецифически влиял на пищевое заглатывание гидр, изменяя как время от захвата пиши до заглатывания, так и время заглатывания пищи. Ни в одном из опытов условной пищевой реакции в ответ на изолированное действие света не наблюдалось.
Во второй серии опытов условным сигналом служила вибрация частотой 60 Гц с длительностью импульса 3—6 с, безусловным — постоянный электрический ток (15 В), подаваемый в виде импульсов длительностью 0,5—3 с. Критерием выработки условного рефлекса была реакция сокращения тела при допороговой вибрации. Для исключения суммации разномодальных раздражителей интервал между сочетаниями был увеличен до 3 мин. Увеличение числа сочетаний до 100 — 200 не привело к снижению порога вибрации.
В третьей серии опытов (Тушмалова, Устинова, 1978, 1979) была изучена возможность выработки химического оборонительного условного рефлекса на свет у гидр. Опыты проводились с животными в возрасте 8 — 10 дней, считая его началом отделение гидры от материнской особи. Условным раздражителем служил свет (1000 лк), безусловным — 0,2 %-ный раствор хлорида натрия (эта концентрация вызывает сокращение всего тела гидры). Условный раздражитель действовал изолированно 30 с, а 30 с его действие совпало с безусловным. Раздражители предъявлялись с интервалом 4 мин, что по данным Л. М. Чайлахяна исключало суммационный эффект. Ежедневно каждая гидра получила 20 сочетаний условного и безусловного раздражителей. В процессе опытов у 100 % гидр наблюдалось уменьшение латентного периода реакции сокращения тела гидры в ответ на безусловный раздражитель с достоверностью Р < 0,001. Однако число условных ответов к 6-му дню опытов увеличилось только до 18,6 %. Следовательно, и в этой серии экспериментов выработка условных рефлексов также не наблюдалась, а была лишь отмечена модификация поведения в виде уменьшения латентного периода реакции на безусловный раздражитель.
Таким образом, результаты опытов, проведенных с различными методическими приемами, оказались аналогичными — они не выявили свойств приобретенных реакций, которые можно было бы определить как условнорефлекторные.
Особенности привыкания кишечнополостных. Исследование закономерностей формирования привыкания кишечнополостных началось относительно недавно. Так, Рушфор (Rushforth, 1963) изучал привыкание гидр к сфокусированному пучку света. Скорость привыкания к свету, подаваемому с частотой 1 — 5 раз в 1 мин, зависела от силы освещенности: в то время как для первого отсутствия ответа при сильном свете понадобилось 40 сокращений, при слабом — всего 3 — 5. В другой группе экспериментов изучалось привыкание к механическим раздражителям: один характеризовался частотой 105 Гц длительностью 2 с и подавался с интервалом в 16 с, другой характеризовался частотой 50 Гц, длительностью 1 с и подавался с интервалом в 19 с. Привыкание к механическому стимулу вырабатывалось тем быстрее, чем чаще подавался сигнал: при частоте 105 Гц — за 8 ч, при частоте 50 Гц — за 20 ч. В этих опытах гидры, "привыкшие" к механическому раздражению, отвечали сокращением на действие света. После удаления щупалец у гидр исчезали сокращения на механические раздражения, но сохранялись реакции на свет. Одновременное же удаление щупалец и гипостома (прикрепительное приспособление в области ротового отверстия) прекращало сокращения на оба названные раздражения. Принципиальная важность этих опытов заключается прежде всего в том, что они доказали существование у гидр различных рецепторов для восприятия света и механического раздражения. Полученные данные согласуются с выводами Лентса (Lentz, 1966) о том, что у гидр рецепторы, воспринимающие свет, локализованы в гипостоме, а воспринимающие механические раздражения — в щупальцах.
Привыкание у гидр может вырабатываться и на химические раздражения (Rushforth, 1965, 1967). На основании анализа экспериментальных данных Рушфорт (Rushforth, 1967) дает следующие критерии привыкания гидр: 1) достаточно частое предъявление стимулов приводит к уменьшению ответа вплоть до прекращения; 2) при прекращении раздражения ответные реакции восстанавливаются; 3) более быстрое развитие привыкания наблюдается при меньшем межстимульном интервале; 4) привыкание развивается тем быстрее, чем слабее стимул; 5) при выработке привыкания к одному из раздражителей сохраняется реакция на сигнал другой модальности.
Результаты привыкания у гидр показали необходимость дальнейшей работы в этом направлении, так как до сих пор отдельные свойства привыкания остались неизученными. В частности, не проводились тесты на растормаживание, недостаточно изучено такое свойство этой реакции, как тренированность. Изучение способности к тренированности затруднено еще и тем, что она может проявляться не только в ускорении выработки реакции, а например, в изменении соотношения отдельных фаз при выработке реакции. Выяснению этих вопросов и были посвящены эксперименты Н. А. Тушмаловой и сотрудников на гидре (Hydra attenuate) (Тушмалова и др., 1975; Бресткина и др., 1978). Критерием выработки привыкания служило прекращение сокращения щупалец при действии пороговых вибрационных импульсов частотой 60 Гц, следующих через каждые 10, 15 и 30 с на протяжении 30 мин. Как и у инфузорий, угашение реакции сокращения щупалец зависело от частоты раздражения: при более частом оно было быстрее. Приобретенная реакция сохранялась на протяжении 15, 60 мин после прекращения раздражения. "Привыкшая" гидра сохраняла реакцию сокращения щупалец в ответ на действие раздражителей большей силы или иной модальности. Как и в опытах с инфузориями, выработка привыкания у гидр зависела от исходного функционального состояния животных. Так, привыкание наиболее быстро вырабатывалось при уровне пищевой возбудимости, наблюдаемой через 4 — 5 дней после кормления животных. Скорость выработки привыкания зависела от возраста гидр — привыкание вырабатывалось быстрее у однодневных гидр по сравнению с 15-дневными (рис. 2).
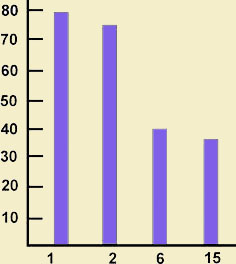
Рис. 2. Динамика выработки привыкания у гидр разного возраста (по: Бресткина и др., 1978) по оси ординат — число ответных реакций %; по оси абсцисс — возраст, сут
Воздействие сильными (намного превышающими пороговый) раздражителями на "привыкшую" гидру не изменяет дальнейшего протекания процесса привыкания. Этот факт может свидетельствовать в пользу того, что, по-видимому, для гидры, так же как и для простейших, нехарактерно явление растормаживания в его типичном проявлении. Гидр повторно через различные промежутки времени подвергали действию механических (вибрационных) раздражителей в режиме привыкания на протяжении 30 мин. В результате выяснилось, что при интервалах, сравнимых со временем сохранения следа после однократного привыкания, наблюдается ярко выраженное ускорение привыкания от опыта к опыту. Однако при интервалах между опытами в 24 ч декремента не наблюдалось. Этот результат представляет интерес, поскольку именно по степени выраженности тренированности у животных различного уровня филогенеза (или нейронных систем различной сложности) при различных интервалах между приложениями можно судить о степени консолидации следа и, возможно, определить четкий критерий различия памяти кратковременной и долговременной — основных функциональных механизмов поведения.
Таким образом прогресс в развитии функциональных механизмов поведения у кишечнополостных по сравнению с простейшими заключается в появлении нового свойства привыкания — тренированности.
Поведение червей
Характеристика условнорефлекторной деятельности плоских червей (планарий). Прогресс в развитии нервной системы на уровне низших (ресничных) червей, или турбеллярий, по сравнению с ранее описанными группами животных выражен началом процесса цефализации. Интегрирующее значение нервной системы на этой стадии филогенеза выражается в регулировании церебральным ганглием важных функций организма, например, координации различных двигательных реакций (Bullock, Horridge, 1965). Вопрос о выработке истинных классических условных рефлексов у низших (ресничных) червей, представителями которых являются планарии, долгое время считался дискуссионным.
Впервые схема периодического сочетания условного раздражителя с безусловным была использована Мак-Коннелом и соавторами (Tompson. McConnel, 1955; McConnel et al., 1959, I960). Эксперименты проводились в специальной камере, снабженной латунными электродами. Действие условного сигнала (свет) на 2 с предшествовало безусловному электрическому раздражению. В течение одного опыта животные получали 150 сочетаний света с электрическим током через каждые 20 с. Критерием выработки условных рефлексов служило сокращение животных в ответ на свет. По окончании серии сочетаний свет — ток частота ответов на свет в виде сокращений и поворотов постоянно повышалась. В контрольных экспериментах, где животные получали избирательно свет или электрический ток (или не подвергались воздействию раздражителей), отмечено лишь небольшое изменение числа ответов. В результате статистической обработки результатов исследования авторы пришли к выводу о возможности выработки классических условных рефлексов у планарий. Наварра (Navarra, 1961) на планарии Dugesia tigrina в течение одного опыта 300 раз сочетал электрическое раздражение со светом и получил аналогичные данные. Ли (Li, 1963), работая с планарией Cura forlmani, в условиях свободного передвижения в качестве подкрепления применял выключение верхнего света при пересечении животными узкого пучка света, направленного параллельно дну экспериментального цилиндра (условный сигнал). У экспериментальных животных по сравнению с контрольными наблюдался большой процент положительных реакций. Вест (Best, 1962) в опытах с планарией Dugesia dorotocephala и Cura forlmani в сложном лабиринте при использовании в качестве условного сигнала света, а безусловного — лишение планарии воды подтвердил способность планарии к дрессировке. В работе Гриффард (Griffard, 1963) с Phagoata gracilis условным раздражителем служил ток воды, безусловным — пропускание электрического тока. На основании проведенных опытов автор также приходит к положительному заключению о выработке условных рефлексов у планарии. Необходимо отметить, что Гриффард впервые проводил эксперименты более длительное время, чем предыдущие авторы, проверяя сохранение условной реакции через 10 — 15 ч.
Используя методику Томпсона и Мак-Коннела (Tompson, McConnel, 1955), Джекобсон и со
|




