

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Тема: Основні форми поведінки. Інстинкт та навчання.
Практичне заняття 2.
Тема: Основні форми поведінки. Інстинкт та навчання.
Мета: Ознайомлення з основними принципами класифікації поведінки тварин; вивчення особливостей вродженої та набутої поведінки, співвідношення вроджених стереотипів, індивідуального та соціального досвіду в поведінці тварин; вивчення складних моделей вродженої (інстинктивної) поведінки тварин, принципів та підходів до її аналізу (К.Лоренц, Н.Тінберген); класифікації і характеристики основних видів навчання у тварин.
Форма заняття: семінар.
Обладнання: методичні рекомендації, навчальні посібники.
План заняття
1. Загальна характеристика видоспецифічної поведінки. Таксиси і тропізми.
- Ж.Леб. Тропізми – елементи поведінки тварин.
2. Рефлекси.
2.1. Класичні умовні рефлекси. Утворення умовних рефлексів.
2.2. Інструментальне навчання.
2.3. Оперантна поведінка.
3. Інстинктивна поведінка як основа життєдіяльності тварин.
3.1. Внутрішні та зовнішні фактори інстинктивної поведінки.
3.2. Релізери та їх властивості. Характеристика «комплексів фіксованих дій» за К.Лоренцом. Гідравлічна модель інстинктивної діяльності.
3.3. Відкриття імпринтингу. Чутливі періоди для зафіксування.
3.4. Ієрархічна модель інстинктивної поведінки за Н. Тінбергеном.
4. Научіння тварин.
4.1. Загальна характеристика процесу научіння.
4.2. Біологічні аспекти навчання. Еволюційні аспекти навчання. Навчання уникнення ворогів.
4.3. Найпростіші форми навчання – габітуація і сенситизація, сенсибілізація.
4.4. Облігатне і факультативне навчання.
4.5. Опосередковане навчання (приклади опосередкованого навчання в роботі Б.П.Мантейфель «Опосредованное обучение»/ Додаток 1) та його біологічне значення.
4.6. Навички і методи їх вивчення.
4.7. Форми навчання, які не пов’язані з підкріпленням.
4.8. Дресирування.
4.9. Навчання та спілкування. Наслідування.
4.10. Закономірності навчання у представників різних систематичних груп.
- Стаття: Тушмалова Н.А. Основные закономерности эволюции поведения беспозвоночных. (Додаток 3).
Завдання для самостійної роботи по підготовці до заняття:
1. Опрацювати літературу за питаннями плану семінару.
2. Підготувати короткий конспект-відповідь на питання семінару.
3. Порівняльна характеристика понять «інстинкт», «рефлекс» та «розум» в роботі А.И.Северцова «Эволюция психики» (Додаток 2).
4. Скласти словник термінів: безумовний рефлекс, умовний рефлекс, імпринтинг, габітуація, навичка, облігатне навчання, факультативне навчання.
Література.
1. Правоторов Г.В. Зоопсихология для гуманитариев. М.: 2005, - Ст. 36, 40, 45 - 64, 81 -104, 281 – 301.
2. Фабри К.Э. Основы зоопсихологии. М.: Изд-во Моск. ун-та, 1993. –
Ст. 41 – 87.
3. Мак-Фарленд Д. Поведение животных. Психобиология, этология и эволюция. М.: Мир, 1988. Ст. 324 – 341.
4. Дьюсбери Д. Поведение животных. М.: Мир, 1981. Ст.182 -199.
- З.А.Зорина, И.И.Полетаева. Элементарное мышление животных. М.: Аспектпресс. 2003 г. – c. 64 – 105.
- Меннинг О. Поведение животных. Вводный курс: Пер. с англ./Перевод Зориной 3. А., Полетаевой И. И.; Под ред. и с предисл. Л. В. Крушинского.— М.: Мир, 1 982,—360 с.
- Зорина З.А., Полетаева И.И., Резникова Ж.И. Основы этологии и генетики поведения
- Шовен, Р. Поведение животных / Р. Шовен; пер. с фр. Л. С. Бондарчука, З. А. Зориной. - Изд. 2-е. - М.:, 2009. - 487 с.
Теми рефератів
1. Імпринтинг та його роль в формуванні різних форм поведінки.
2. Опосередковане навчання та його роль в формуванні поведінки.
3. Фізіологічні механізми розвитку інстинктивної поведінки.
4. Основні методи вивчення інстинктів.
5. Неасоціативне навчання і його роль в формуванні поведінки.
6. Формування поведінки тварин за Л.В. Крушинським.
7. Ф. Скіннер та його вклад в науку про поведінку.
Додаток 1
Додаток 3
Додаток 3
Поведение кишечнополостных
Об условнорефлекторной деятельности кишечнополостных. Вопрос о способности кишечнополостных формировать условные рефлексы в настоящее время остается открытым. Результаты немногочисленных экспериментальных попыток выработки условных реакций у различных видов этого типа до сих пор не дали положительных результатов. Одна из первых работ была предпринята А. А. Зубковым и Г. Г. Поликарповым (1951). Они проводили опыты в часовом стекле и обнаружили, что движения гидры, прикрепленной близко к поверхности воды, совершаются в направлении от поверхности воды, хотя на глубине в условиях равномерного освещения наблюдаются движения во всех направлениях; через 60 мин после начала опыта движения в сторону поверхности воды прекращаются. При вторичном повышении уровня воды "выработанная" реакция сохраняется до 3—4 ч, а затем угасает. На основании таких свойств реакции, как угашение при неподкреплении, торможение при действии экстрараздражителей (сотрясение и изменение освещенности), авторы определили наблюдаемую модификацию поведения гидр как выработку условного рефлекса, однако результаты проведенных опытов могут иметь и иные объяснения. Так, Л. М. Чайлахян (1957) объясняет "приобретенность" такой реакции гидр лишь изменением состояния мышечных элементов. Более детальный анализ условнорефлекторной деятельности кишечнополостных дан в работе Л. М. Чайлахяна (1957) на примере опытов с пресноводным полипом. Сочетание света (условный сигнал длительностью от 5 с до 1 мин) с электрическим током — безусловный раздражитель при интервалах от 2 до 3 мин привело к проявлению реакции сокращения животных только на изолированное действие света. При увеличении интервала между условными и безусловными раздражителями до 5—6 мин "условная" реакция отсутствовала. Выявленные закономерности позволили автору заключить, что реакция полипа на свет не условный, а суммационный рефлекс — результат повышения возбудимости животного вследствие суммации следов от предыдущих раздражений электрическим током, под влиянием чего допороговая интенсивность света становилась пороговой.
Нами была проведена серия экспериментов по изучению способности к формированию условнорефлекторной памяти у пресноводной гидры (Hydra attenuate). В первых двух сериях опытов (Тушмалова, Симирский, 1977) были исследованы реакции гидр на действие сочетанных раздражителей при попытке выработать пищевые и электрооборонительные условные рефлексы. Условным сигналом служил свет лампы освещенностью в 1000 лк с изолированным действием в течение 30—60 с, на протяжении 30 с совпадающий с безусловным пищевым подкреплением (циклопы). Интервалы между сочетаниями определялись временем заглатывания циклопа и составляли 6 — 10 мин. Критерием положительной условной реакции было выбрано открывание гипостома в ответ на изолированное действие света. Контролем служили животные, которые ежедневно получали дозированную пищу в отсутствии света, и животные, получающие пишу при непрерывном освещении красным светом. Изменение реакции гидр от сочетания к сочетанию выражалось лишь в том, что свет неспецифически влиял на пищевое заглатывание гидр, изменяя как время от захвата пиши до заглатывания, так и время заглатывания пищи. Ни в одном из опытов условной пищевой реакции в ответ на изолированное действие света не наблюдалось.
Во второй серии опытов условным сигналом служила вибрация частотой 60 Гц с длительностью импульса 3—6 с, безусловным — постоянный электрический ток (15 В), подаваемый в виде импульсов длительностью 0,5—3 с. Критерием выработки условного рефлекса была реакция сокращения тела при допороговой вибрации. Для исключения суммации разномодальных раздражителей интервал между сочетаниями был увеличен до 3 мин. Увеличение числа сочетаний до 100 — 200 не привело к снижению порога вибрации.
В третьей серии опытов (Тушмалова, Устинова, 1978, 1979) была изучена возможность выработки химического оборонительного условного рефлекса на свет у гидр. Опыты проводились с животными в возрасте 8 — 10 дней, считая его началом отделение гидры от материнской особи. Условным раздражителем служил свет (1000 лк), безусловным — 0,2 %-ный раствор хлорида натрия (эта концентрация вызывает сокращение всего тела гидры). Условный раздражитель действовал изолированно 30 с, а 30 с его действие совпало с безусловным. Раздражители предъявлялись с интервалом 4 мин, что по данным Л. М. Чайлахяна исключало суммационный эффект. Ежедневно каждая гидра получила 20 сочетаний условного и безусловного раздражителей. В процессе опытов у 100 % гидр наблюдалось уменьшение латентного периода реакции сокращения тела гидры в ответ на безусловный раздражитель с достоверностью Р < 0,001. Однако число условных ответов к 6-му дню опытов увеличилось только до 18,6 %. Следовательно, и в этой серии экспериментов выработка условных рефлексов также не наблюдалась, а была лишь отмечена модификация поведения в виде уменьшения латентного периода реакции на безусловный раздражитель.
Таким образом, результаты опытов, проведенных с различными методическими приемами, оказались аналогичными — они не выявили свойств приобретенных реакций, которые можно было бы определить как условнорефлекторные.
Особенности привыкания кишечнополостных. Исследование закономерностей формирования привыкания кишечнополостных началось относительно недавно. Так, Рушфор (Rushforth, 1963) изучал привыкание гидр к сфокусированному пучку света. Скорость привыкания к свету, подаваемому с частотой 1 — 5 раз в 1 мин, зависела от силы освещенности: в то время как для первого отсутствия ответа при сильном свете понадобилось 40 сокращений, при слабом — всего 3 — 5. В другой группе экспериментов изучалось привыкание к механическим раздражителям: один характеризовался частотой 105 Гц длительностью 2 с и подавался с интервалом в 16 с, другой характеризовался частотой 50 Гц, длительностью 1 с и подавался с интервалом в 19 с. Привыкание к механическому стимулу вырабатывалось тем быстрее, чем чаще подавался сигнал: при частоте 105 Гц — за 8 ч, при частоте 50 Гц — за 20 ч. В этих опытах гидры, "привыкшие" к механическому раздражению, отвечали сокращением на действие света. После удаления щупалец у гидр исчезали сокращения на механические раздражения, но сохранялись реакции на свет. Одновременное же удаление щупалец и гипостома (прикрепительное приспособление в области ротового отверстия) прекращало сокращения на оба названные раздражения. Принципиальная важность этих опытов заключается прежде всего в том, что они доказали существование у гидр различных рецепторов для восприятия света и механического раздражения. Полученные данные согласуются с выводами Лентса (Lentz, 1966) о том, что у гидр рецепторы, воспринимающие свет, локализованы в гипостоме, а воспринимающие механические раздражения — в щупальцах.
Привыкание у гидр может вырабатываться и на химические раздражения (Rushforth, 1965, 1967). На основании анализа экспериментальных данных Рушфорт (Rushforth, 1967) дает следующие критерии привыкания гидр: 1) достаточно частое предъявление стимулов приводит к уменьшению ответа вплоть до прекращения; 2) при прекращении раздражения ответные реакции восстанавливаются; 3) более быстрое развитие привыкания наблюдается при меньшем межстимульном интервале; 4) привыкание развивается тем быстрее, чем слабее стимул; 5) при выработке привыкания к одному из раздражителей сохраняется реакция на сигнал другой модальности.
Результаты привыкания у гидр показали необходимость дальнейшей работы в этом направлении, так как до сих пор отдельные свойства привыкания остались неизученными. В частности, не проводились тесты на растормаживание, недостаточно изучено такое свойство этой реакции, как тренированность. Изучение способности к тренированности затруднено еще и тем, что она может проявляться не только в ускорении выработки реакции, а например, в изменении соотношения отдельных фаз при выработке реакции. Выяснению этих вопросов и были посвящены эксперименты Н. А. Тушмаловой и сотрудников на гидре (Hydra attenuate) (Тушмалова и др., 1975; Бресткина и др., 1978). Критерием выработки привыкания служило прекращение сокращения щупалец при действии пороговых вибрационных импульсов частотой 60 Гц, следующих через каждые 10, 15 и 30 с на протяжении 30 мин. Как и у инфузорий, угашение реакции сокращения щупалец зависело от частоты раздражения: при более частом оно было быстрее. Приобретенная реакция сохранялась на протяжении 15, 60 мин после прекращения раздражения. "Привыкшая" гидра сохраняла реакцию сокращения щупалец в ответ на действие раздражителей большей силы или иной модальности. Как и в опытах с инфузориями, выработка привыкания у гидр зависела от исходного функционального состояния животных. Так, привыкание наиболее быстро вырабатывалось при уровне пищевой возбудимости, наблюдаемой через 4 — 5 дней после кормления животных. Скорость выработки привыкания зависела от возраста гидр — привыкание вырабатывалось быстрее у однодневных гидр по сравнению с 15-дневными (рис. 2).
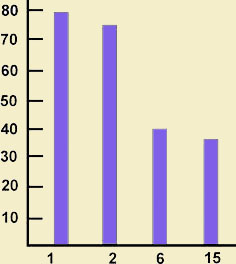
Рис. 2. Динамика выработки привыкания у гидр разного возраста (по: Бресткина и др., 1978) по оси ординат — число ответных реакций %; по оси абсцисс — возраст, сут
Воздействие сильными (намного превышающими пороговый) раздражителями на "привыкшую" гидру не изменяет дальнейшего протекания процесса привыкания. Этот факт может свидетельствовать в пользу того, что, по-видимому, для гидры, так же как и для простейших, нехарактерно явление растормаживания в его типичном проявлении. Гидр повторно через различные промежутки времени подвергали действию механических (вибрационных) раздражителей в режиме привыкания на протяжении 30 мин. В результате выяснилось, что при интервалах, сравнимых со временем сохранения следа после однократного привыкания, наблюдается ярко выраженное ускорение привыкания от опыта к опыту. Однако при интервалах между опытами в 24 ч декремента не наблюдалось. Этот результат представляет интерес, поскольку именно по степени выраженности тренированности у животных различного уровня филогенеза (или нейронных систем различной сложности) при различных интервалах между приложениями можно судить о степени консолидации следа и, возможно, определить четкий критерий различия памяти кратковременной и долговременной — основных функциональных механизмов поведения.
Таким образом прогресс в развитии функциональных механизмов поведения у кишечнополостных по сравнению с простейшими заключается в появлении нового свойства привыкания — тренированности.
Поведение червей
Характеристика условнорефлекторной деятельности плоских червей (планарий). Прогресс в развитии нервной системы на уровне низших (ресничных) червей, или турбеллярий, по сравнению с ранее описанными группами животных выражен началом процесса цефализации. Интегрирующее значение нервной системы на этой стадии филогенеза выражается в регулировании церебральным ганглием важных функций организма, например, координации различных двигательных реакций (Bullock, Horridge, 1965). Вопрос о выработке истинных классических условных рефлексов у низших (ресничных) червей, представителями которых являются планарии, долгое время считался дискуссионным.
Впервые схема периодического сочетания условного раздражителя с безусловным была использована Мак-Коннелом и соавторами (Tompson. McConnel, 1955; McConnel et al., 1959, I960). Эксперименты проводились в специальной камере, снабженной латунными электродами. Действие условного сигнала (свет) на 2 с предшествовало безусловному электрическому раздражению. В течение одного опыта животные получали 150 сочетаний света с электрическим током через каждые 20 с. Критерием выработки условных рефлексов служило сокращение животных в ответ на свет. По окончании серии сочетаний свет — ток частота ответов на свет в виде сокращений и поворотов постоянно повышалась. В контрольных экспериментах, где животные получали избирательно свет или электрический ток (или не подвергались воздействию раздражителей), отмечено лишь небольшое изменение числа ответов. В результате статистической обработки результатов исследования авторы пришли к выводу о возможности выработки классических условных рефлексов у планарий. Наварра (Navarra, 1961) на планарии Dugesia tigrina в течение одного опыта 300 раз сочетал электрическое раздражение со светом и получил аналогичные данные. Ли (Li, 1963), работая с планарией Cura forlmani, в условиях свободного передвижения в качестве подкрепления применял выключение верхнего света при пересечении животными узкого пучка света, направленного параллельно дну экспериментального цилиндра (условный сигнал). У экспериментальных животных по сравнению с контрольными наблюдался большой процент положительных реакций. Вест (Best, 1962) в опытах с планарией Dugesia dorotocephala и Cura forlmani в сложном лабиринте при использовании в качестве условного сигнала света, а безусловного — лишение планарии воды подтвердил способность планарии к дрессировке. В работе Гриффард (Griffard, 1963) с Phagoata gracilis условным раздражителем служил ток воды, безусловным — пропускание электрического тока. На основании проведенных опытов автор также приходит к положительному заключению о выработке условных рефлексов у планарии. Необходимо отметить, что Гриффард впервые проводил эксперименты более длительное время, чем предыдущие авторы, проверяя сохранение условной реакции через 10 — 15 ч.
Используя методику Томпсона и Мак-Коннела (Tompson, McConnel, 1955), Джекобсон и соавторы (Jacobson et al., 1967) также пришли к выводу о возможности выработки у планарий классических условных рефлексов. Эксперименты Томпсона точно воспроизвёл Хэлас с сотрудниками (Halas et al., 1962). Однако эти авторы рассматривают реакции планарии на условный сигнал (свет) как рефлекс сенсибилизации. А. Н. Черкашин и соавторы (1966) изучали реакции планарии на сочетание монофазного электрического тока (безусловный раздражитель) и света (условный раздражитель). Авторы делают заключение о том, что в данных методических условиях приобретенные реакции не достигают уровня условных рефлексов.
Таким образом, вопрос о физиологических механизмах приобретенных реакций у планарий оставался открытым. Для окончательного ответа на него были проведены дальнейшие эксперименты на 5 видах планарий с различной экологией, включая эндемиков озера Байкал. В первой серии вырабатывался электрооборонительный условный рефлекс на свет (Воронин, Тушмалова, 1965). Оказалось, что для первого проявления реакции, сходной с условнорефлекторной, понадобилось от 51 до 76 сочетаний. Однако упрочения ее не происходило, несмотря на 300 сочетаний раздражителей. Во второй серии опытов в качестве условного раздражителя применялся слабый электрический ток, вызывавший ориентировочный рефлекс в виде поворота головного конца тела животного, и в качестве безусловного — отрицательная реакция планарий на свет (уплывание в сторону неосвещенной части желоба). Последовательного закрепления реакции из опыта в опыт, так характерного для классических условных рефлексов, не наблюдалось и в этом случае. Временная связь у всех планарий была нестойкой, количество положительных реакций из каждых 10 сочетаний условного сигнала с безусловным редко превышало 50%.
Была использована и более простая схема экспериментов, в которой условный сигнал был постоянным. Дно экспериментальной камеры было разделено на темную и светлую половины. В опыте сначала определялось время нахождения животных на темном и светлом полях, затем на одном из полей они получали удар электрическим током, вызывающим продольное сокращение тела. Основным критерием выработки условной реакции в этой серии экспериментов служило уменьшение времени нахождения планарий в той части камеры, где возникало электрическое раздражение. Закономерного и стойкого увеличения времени пребывания животных на поле, где животным не наносилось безусловное раздражение, не наблюдалось. В дальнейшем был применен более адекватный безусловный раздражитель — раствор поваренной соли (Тушмалова, Громыко, 1968). В этом случае максимальный уровень положительных реакций не превышал 50 — 70%, и, как при электрическом подкреплении, они характеризовались нестойкостью от опыта к опыту и в течение одного эксперимента. Однако при этом возможно было четко отдифференцировать начало выработки условных рефлексов (после 40 — 60 сочетаний), максимальный уровень положительных реакций (120 — 200 сочетаний) и снижение уровня условнорефлекторной деятельности (сокращение числа положительных реакций от 10 до 0 %) после 200 — 300 сочетаний. У контрольных животных при изолированном применении света в 100% случаев наблюдалось лишь слабое возбуждение, выражающееся в сокращении (не более 8 — 10% по отношению к положительным реакциям опытных животных). Аналогичная картина наблюдалась в опытах с псевдообучением (предъявление условного и безусловного раздражителей в случайном порядке). При остром угашении реакции самопроизвольно не восстанавливались. Из анализа экспериментального материала следует, что условные рефлексы у изученных пресноводных планарий недостаточно стойки, не обладают всеми качествами классических условных рефлексов. Они характеризуются следующими признаками, общими для экологически различных видов животных: непрочностью в течение одного опыта, непрочностью от опыта к опыту (упрочение не наступало даже после 335 сочетаний условного сигнала с безусловным), угасанием реакций после 200 — 300 сочетаний, несмотря на подкрепление. Перечисленные свойства условных рефлексов не являются отражением индивидуальных особенностей отдельных видов, так как они характерны для животных с различной экологией (от живущих в пойменных ручьях Подмосковья до эндемиков Байкала) (Тушмалова, 1968, 1967). Отмеченные закономерности не являются и следствием примененных методических приемов, аналогичные данные были получены во всех 6 сериях опытов, значительно отличающихся методически.
Таким образом, подобные реакции можно отнести к категории лишь примитивных нестойких условных рефлексов, свойственных животным определенного уровня филогенетического развития (Воронин и др., 1972; Тушмалова, 1976).
Особенности строения головного ганглия планарий. Одна из замечательных филогенетических особенностей ресничных червей, к которым относятся планарии, заключается в том, что они представляют уровень "перехода" диффузной нервной сети в концентрированную систему. У ресничных червей впервые в эволюции нервные элементы концентрируются на переднем конце тела (Hanstrom, 1928; Заварзин, 1941; Беклемишев, 1964), т. е. появляются зачатки цефализации. Детальные сведения о клеточном (цитоархитектоническом) строении этого примитивного мозга в литературе долгое время отсутствовали, и лишь стремление определить степень соответствия примитивного поведения планарий структурной организацией их "мозга" послужило поводом к конкретным исследованиям. Строение мозга планарий было изучено на примере типичного представителя ресничных червей P. nigra (Бочарова, Тушмалова, 1968; Свешников и др., 1968; Тушмалова, Свешников, 1972).
Церебральный ганглий P. nigra расположен приблизительно в 500 мкм от головного конца тела и состоит из двух частей. Из-за отсутствия оболочки его границы могут быть определены относительно. Характерной особенностью головного ганглия планарий служит наличие в нем многочисленных специфических образований, состоящих из клеток, которые располагаются вокруг проходящих через мозг дорсовентральных мышц (инсулярные, или островковые включения по: Bohmig. 1906), в количестве от 40 до 70. В состав ганглия входят разнообразные по величине и форме клетки (большинство биполярные, реже мультиполярные). Особенностью большинства клеток является бедность цитоплазмой и отсутствие ядрышек у большинства клеток. Размеры клеток варьируют от 3 — 5, 10 — 15 до 15 — 20 ("гигантские" клетки) мкм. Последние отличаются богатой цитоплазмой, большим ядрышком и высоким содержанием РНК в цитоплазме. Наличие в цитоплазме этих клеток характерной зернистой субстанции дало право считать их наиболее дифференцированными нейронами всего мозга.
Таким образом, исследования показали, что церебральный ганглий планарий несет черты примитивной организации, состоит в основном из мелких, малодифференцированных клеток. Основной особенностью гистологического строения ганглия служит наличие в нем большого количества мышечных и паренхиматозных клеток, выполняющих, по-видимому, нейросекреторные функции. Только отдельные ("гигантские") клетки могут быть с большей степенью уверенности классифицированы как нервные.
Представление о примитивности нервной системы планарий разделяют многие авторы (Кричинская, 1972; Лурье, 1975). Они считают, что все нервные клетки планарий бифункциональны и обладают способностью к нейросекреции. Таким образом, появившаяся впервые в эволюции у планарий цефализация представлена здесь в своей примитивной форме и к ней приурочена приобретаемая в индивидуальной жизни организма примитивная форма нестойких условных рефлексов.
Свойства условнорефлекторной деятельности кольчатых червей. Среди большой группы кольчатых червей, являющихся эволюционными потомками плоских червей, особое место занимают представители класса олигохет — дождевые черви, на которых проводились основные опыты по условным рефлексам. Наиболее подробно способность к формированию условных рефлексов у дождевых червей Lumbricus rubellus изучена Н. М. Хоничевой (1968). Она вырабатывала оборонительные условные реакции в Т-образном лабиринте. Черви обучались поворачивать в правый или левый рукав лабиринта. Безусловным раздражителем служил переменный ток различной интенсивности, а условным — сам лабиринт, элементы которого, вероятно, воспринимались проприоцептивной и тактильной афферентациями. Критерием выработки рефлекса служило увеличение числа по воротов в рукав лабиринта, где животные не подвергались электрической стимуляции. Максимальное число сочетаний доходило до 400. Однако постепенного, закономерного увеличения числа положительных реакций отмечено не было. Характерной особенностью обучения червей явилось колебание процента положительных реакций.
Сравнение динамики обучения трех групп червей показало, что повышение силы безусловного раздражителя (электрического тока) увеличивает скорость выработки реакций. Так, при напряжении 5 В максимальный процент положительных реакций был отмечен на 20-й опытный день, при 12 В — на 17-й опытны день, при 18 — уже на 11-й день. Оказалось, что эта тенденция к более быстрому обучению осуществляется за счет сокращения продолжительности его 1-й стадии, которая равнялась 12, 8 и 5 дням соответственно для слабого, среднего и сильного подкрепляющего раздражителя. Наблюдалась зависимость максимального процента положительных реакций от величины подкрепляющего раздражителя: при 5 В — 54, при 12 В — 70, при 18 В — 85%. Существенно, что удаление надглоточного ганглия не изменяло динамику обучения — факт, несомненно свидетельствующий в пользу незначительной интегрирующей роли данного отдела мозга.
Совсем иные закономерности были получены в опытах на полихетах. Так, Копеланд (Gopeland, 1930) у Nereis virens выработал устойчивый условный рефлекс на тактильное раздражение. У Nereis pelagica (беломорская форма) была четко продемонстрирована возможность выработки пищевых условных рефлексов на свет и вибрацию (Воронин и др., 1982). Критерием выработки условных рефлексов служило изменение "знака" фотонегативной реакции. До начала опытов черви постоянно находились внутри стеклянной трубки, а после 30 — 50 сочетаний условного раздражителя с пищей животные проявляли четкую поисковую пищевую реакцию, выходя из трубки. Анализ результатов показал, что у полихет вырабатываются реакции, обладающие всеми основными свойствами истинных условных рефлексов: возрастание числа положительных ответов от опыта к опыту, высокий максимальный процент положительных реакций (до 80 — 100) и длительность их сохранения (до 6 — 15 дней). Весьма существенно, что выработанная реакция угасала при отсутствии подкрепления и самопроизвольно восстанавливалась. В контрольных опытах — при псевдообуславливании — увеличения числа положительных ответов не наблюдалось.
Выявленные закономерности условнорефлекторной деятельности полихет коррелируют с относительно дифференцированным мозгом животных. Хорошо известно, что одной из особенностей их мозга является возникновение специального ассоциативного центра — грибовидных тел. Удаление этих отделов мозга приводит к нарушению условных рефлексов, как показано в опытах на пчелах (Воскресенская, 1957). Таким образом, истинные условные рефлексы как один из достаточных совершенных механизмов, определяющих приобретенное поведение, впервые в эволюции, по-видимому, появляются у полихет (Тушмалова, 1980).
Поведение членистоногих
Характерной эволюционной особенностью представителей данной систематической группы служит высокая степень цефализации. Другой отличительной чертой является достаточно четко выраженная структурно-функциональная специализация мозга. Названные особенности наложили отпечаток на свойства функциональных механизмов, определяющих приобретенное поведение, закономерности которого изучаются на чисто поведенческом и нейрофизиологическом уровнях. Сказанное относится прежде всего к насекомым.
Высшая нервная деятельность наиболее детально изучалась у ракообразных и насекомых.
Условные рефлексы ракообразных. Беспозвоночные этой группы резко отличаются по морфологической организации нервной системы: у низших раков она имеет признаки кольчатых червей, что отражается и в функциональных механизмах поведения. Положительные результаты выработки условного рефлекса у дафний были получены лишь в своеобразных опытах А. Б. Когана и А. П. Семенова (1955). У дафний был выработан пищевой условный рефлекс по схеме: кормление в темноте (ночь), голодание на свету (день).
На высших ракообразных были получены результаты при использовании обычной схемы выработки условных реакций. В опытах на раках-отшельниках Сполдинг (Spaulding, 1904) выработал относительно стойкий (сохранялся 7 дней) пищевой условный рефлекс на затемнение. Возможность выработки угасательного и дифференцировочного торможения у рака-отшельника (Pagurus striatus) была продемонстрирована Михайловым (Mikhailoff, 1922) еще в 1922 году. Положительные результаты были получены в опытах А. Я. Карась (1964) на ресничных раках Astacus leptodactilis по выработке двигательного пищевого условного рефлекса на свет. Наиболее правдоподобно закономерности высшей нервной деятельности у высших раков были исследованы в серии работ Л. Я. Карась (1962, 1963, 1964) на крабах Carcinus means. Была показана возможность выработки пищевых, зрительных, тактильных условных рефлексов и условных рефлексов на раздражение статорецепторов и выработки основных видов внутреннего торможения (угасательного, дифференцировочного и условного тормоза). Особенностью положительных и отрицательных условных связей была их нестойкость. Весьма существенно, что у животных этого уровня филогенеза формировались условные рефлексы на цепные и комплексные раздражители. Однако в отличие от насекомых (пчел) эти реакции также отличались нестойкостью. Автор предполагает, что сигнальным признаком цепных раздражителей была сила отдельного компонента, а не их определенная последовательность. Такое представление свидетельствует о низком уровне аналитико-синтетических свойств нервной системы ракообразных. Морфологической основой сформулированного представления может быть существование связей в ЦНС этих животных (Коштоянц, 1957).
Заключение
Анализ работ, посвященных изучению приобретенных форм поведения у беспозвоночных, позволяет сделать заключение о том, что способность к накоплению индивидуального опыта появляется в эволюции на самых ранних ступенях развития — у донервных простейших организмов (Тушмалова, 1976, 1981). Однако функциональные механизмы, определяющие поведение беспозвоночных животных разного уровня филогенеза, различны (Тушмалова, 1980). Так, из двух форм эволюции ВНД (Воронин, 1977) сигнальная форма индивидуального приспособления появляется лишь у полихет, а низшим червям, кишечнополостным и простейшим свойственна несигнальная форма приспособления.
Для рассмотрения основных закономерностей эволюции поведения беспозвоночных принципиальное значение имеет факт формирования приобретенного поведения у донервных организмов за счет макромолекулярных и ультраструктурных перестроек (Тушмалова и др., 1974; Зазулина, 1979). В эволюционном плане весьма существенно, что характер этих перестроек однозначен у животных разного уровня филогенеза, обладающих различными функциональными механизмами приобретенного поведения (Тушмалова, 1980, 1983).
Исходя из представлений Л. А. Орбели (1961) о том, что в процессе эволюционного развития старые функциональные отношения не уничтожаются, а лишь затормаживаются новыми, можно высказать предположение о сохранении протоплазмой клеток организмов, обладающих нервной системой, свойства простейших — способности формировать следовые реакции за счет внутриклеточных перестроек. Развиваемое нами представление согласуется с мыслью П. К. Анохина (1968), считающего, что эволюция изменила лишь "средства доставки информации", но ее конечная обработка осуществляется одним и тем же способом — молекулярным. Следовательно, можно предположить, что усложнение (совершенствование) функциональных механизмов поведения по мере возрастания эволюционного уровня определяется структурными изменениями нервной системы. Этот вывод на примере беспозвоночных иллюстрирует одно из существенных методологических положений нейрофизиологии о зависимости "качественно нового ранга рефлекса — условного рефлекса" от уровня развития нервной систем (Карамян, 1969). Действительно, при сравнении функциональных механизмов поведения беспозвоночных обращает на себя внимание параллель в степени сложности структурной функциональной организации. Это есть не что иное, как выражение морфофизиологического прогресса — ароморфоза по А.С. Северцову (1967) (см. табл. 1).
Таблица 1
Соотношение морфологических и функциональных ароморфозов на примере эволюции поведения беспозвоночных
| Группы животных
| Ароморфозы
| |
| Морфологический уровень (организация нервной системы)
| Функциональный уровень (высшая форма физиологических механизмов поведения)
| | Простейшие
| Донервный уровень
| Привыкание
| | Кишечнополостные
| Примитивная нервная система
| Привыкание + тренированность
| | Плоские черви
| Зачатки цефализации
| Нестойкие условные рефлексы
| | Кольчатые черви
| Ассоциативный центр мозга — грибовидные тела
| Истинные условные рефлексы
| | Насекомые
| Структурно-функциональльная дифференцировка мозга
| Продолжительные и отрицательные условные рефлексы, инсnрументальные условные рефлексы, цепные условные рефлексы.
| Таким образом, основные закономерности функциональных механизмов поведения беспозвоночных являются по существу филогенетическим отражением принципа единства структуры и функции.
Практичне заняття 2.
Тема: Основні форми поведінки. Інстинкт та навчання.
Мета: Ознайомлення з основними принципами класифікації поведінки тварин; вивчення особливостей вродженої та набутої поведінки, співвідношення вроджених стереотипів, індивідуального та соціального досвіду в поведінці тварин; вивчення складних моделей вродженої (інстинктивної) поведінки тварин, принципів та підходів до її аналізу (К.Лоренц, Н.Тінберген); класифікації і характеристики основних видів навчання у тварин.
Форма заняття: семінар.
Обладнання:<
|




