Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Потенциалы поляризации мембран клеток.Содержание книги
Поиск на нашем сайте
Открытое Гальвани (1770е годы) «живое» электричество почти два века не было описано в общих понятиях теории электричества. «Химическое» электричество Волта открыто в те же годы, а Фарадей описал свои эксперименты по связи между магнитным и электрическим полем в 1831г. В этом же году по Неве двигался катер на электромоторе, через 15 лет в Европе ходил трамвай. Только в 1889г В.Г. Нернст (1864-1941) опубликовал уравнения, определяющие электро химический потенциал электрода. Он же предложил схему «концентрационного элемента», разделенного полупроницаемой мембраной (эквивалент строения клетки). Был рассчитан потенциал поляризации мембраны этого элемента (приблизительно 80мВ. Нобелевская премия 1920г.). Окончательно теория поляризации и деполяризации мембран клеток (и экспериментальная проверка) была опубликована в 1959г. А.Л.Ходжкиным (1914-1998. Нобелевская премия 1963г.). Потенциал поляризации клетки является вторым электро биологическим сигналом после тока канала мембраны. Потенциал клетки определяется 1) диффузионными процессами в электролитах, 2) свойствами полупроницаемой мембраны и 3) особыми процессами переноса ионов К+ и Na+ через мембрану (калий- натриевый насос). Диффузионное движение ионов в электролите определяется законом Фика: Jd = - D dC/dx, где Jd – число ионов, переносимых через единичную площадку в секунду, С – концентрация ионов, D- коэффициент диффузии, (Для газа D=LV/3, где L – средняя длина пробега, V – средняя скорость. Но D обычно определяется экспериментально). Коэффициент диффузии D определяется через уравнение Эйнштейна (1879-1953): Dp = upRT/F|Zp|, Dp коэффициент диффузии, up – подвижность, R – газовая постоянная 8,314 Дж/ моль при 270С, T – абсолютная температура Кельвина, Zp – валентность, F – число Фарадея 96,487 Кл/гэкв, (RT / F = 23,8 мВ при 270С). На ионы, кроме диффузии, воздействует электрическое поле. Ионный ток в этом случае: Je = - up[Zp/|Zp|]CpE, где Je – плотность ионного тока в молях на ед. площади в сек., up – подвижность иона (скорость при Е=1), р – тип иона. Zp – валентность иона, Zp /| Zp | - направление силы (| Zp |- модуль), Cp – концентрация ионов р, E - напряженность поля (Е=dФ/dx, Ф – поле электрического потенциала). Суммарная плотность тока Jp ионов типа р равна: Jp = Jd + Je = - Dp [ dCp / dx + EZpCpF / RT ]. Клетка охвачена полупроницаемой мембраной. Внутри ионы q, снаружи - p. Для р мембрана проницаема в обе стороны, для q – нет. Возникают диффузионный и электронный токи. При равновесии ток равен нулю. Jp = 0= - Dp [ dCp / dx + EZpCpF / RT ]. (Это равновесие Нернста, 1889г). Нам удобнее Е заменить через dФ/dx. Тогда dCp / dx = - d Ф/ dx [ ZpCpF / RT ]. После разделения переменных, dCp / Cp = - [ ZpF / RT ] d Ф. Интегрируем правую и левую части в пределах от внешнего пространства е до внутреннего i. Интеграл от dФ равен приращению Ф между е и i (т.е. разности потенциалов Vei) Vei = (58/Zp) lg[ [ Cp ]i / [ Cp ]e ] мВ. Применим это выражение к клетке аксона кальмара, концентрации ионов у которого приведены в таблице: Таблица. Ионная концентрация моль на литр.
Подставляя значения Cp e и Cp i для К+ получаем Ек= - 58 lg [397/20]= - 74,7 мВ.
Когда клетка отгорожена от внешнего мира поляризованной мембраной, то поступление питательных веществ отсутствует. Самостийно (примерно раз в 2 секунды или чаще, под воздействием внешних стимулов) клетки деполяризуются. Поляризация исчезает. Клетки на 100 – 200мс. общаются с внешним миром. Мы наблюдаем импульсы деполяризации См рис 4.4.
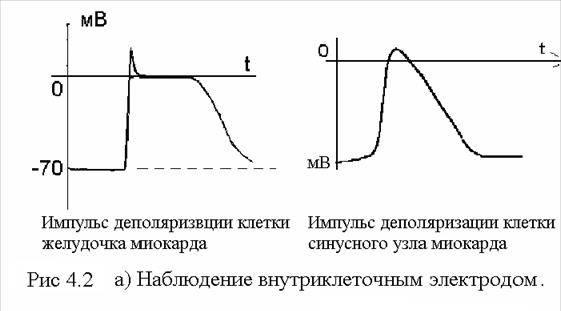
На рис 4.5 показан процесс возбуждения клетки пороговыми импульсами тока: поведение потенциала позволяет делать заключение о сложности происходящих процессов. Релаксационное начало деполяризации происходит с задержкой порядка 0, 2 мс после окончания воздействия стимула. 4.4. Наблюдение деполяризации клетки внутренним и внешним электродом. Следует подчеркнуть, что форма сигнала деполяризации резко отличается при наблюдении внутриклеточным и внешними электродами. На рис 4.4 электрод А введен внутрь клетки. Начальная разность потенциалов (порядка -70 мВ) в момент деполяризации резко спадает (длительность около 1 мс). При этом формируется положительный выброс до 16 мВ. Потенциал переходит в нулевое плоское плато и примерно через 200мс (для клеток миокарда) начинается процесс реполяризации: мембрана восстанавливает свои избирательные свойства для ионов и медленно (за время порядка 100 мс) возрождает потенциал -70 мВ. Эта форма сигнала наблюдается внутриклеточным электродом.
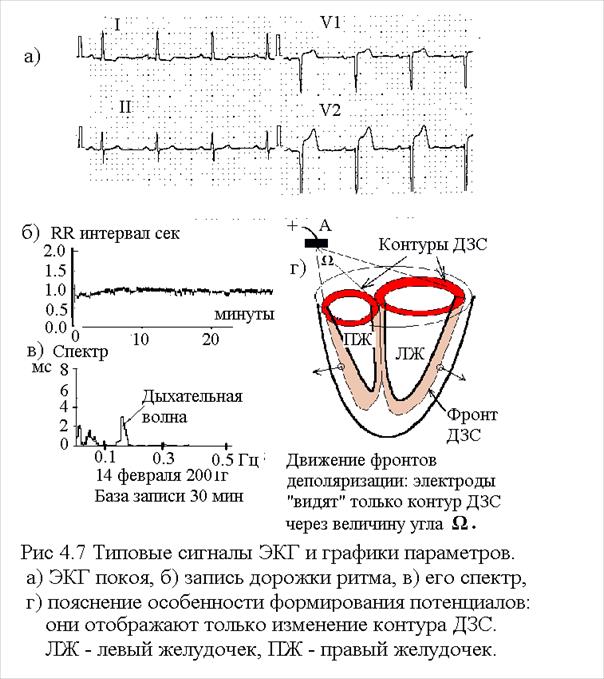
4.5. Сигнал деполяризации длинного нейрона. Возбуждение клетки переходит в длинный отросток эфферентного нейрона. Длина достигает одного метра и более. Процесс деполяризации распространяется со скоростью около 60 мс, передний фронт имеет резкую границу (бегущую вдоль нейрона). Найдем форму сигнала наблюдаемую внешними электродами. Пусть регистрирующий электрод расположен в точке А вне клетки. Сконструируем искусственный элемент (назовем его блок), состоящий из двух ДЗС, расположенных очень близко и встречно друг к другу (см рис 4.7). Внешнее поле такого блока равно нулю, ибо встречные ДЗС компенсируют друг друга. Следовательно этот блок можно перемещать произвольно в пространстве, не нарушая общую картину электрического поля. Расположим наш блок в области бегущего фронта деполяризации нейрона. Один из двух ДЗС нашего блока "замкнет" ДЗС оставшейся не деполяризованной части нейрона. В результате этот замкнутый нейрон не будет иметь внешнего поля. Останется поле второй части нашего блока: этот ДЗС и будет отражать реальную картину поля фронта деполяризующегося нейрона. Он "бежит" вдоль нейрона со скоростью деполяризации.
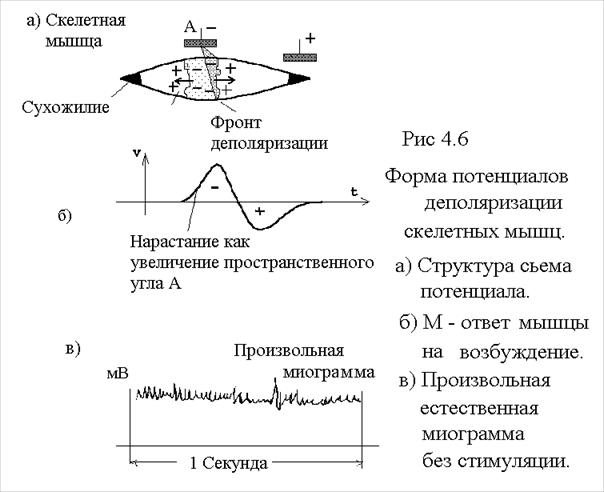
4.6. Сигнал деполяризации скелетных мышц (Рис 4.8)
4.7. Сигналы деполяризации миокарда (рис 4.9) Мышца сердца (миокард) не разделена на отдельные "двигательные единицы", она деполяризуются автономно как "сцинтилий" (вспышка). Управляющие нейроны высшей нервной системы регулируют только частоту возбуждения - ЧСС (пульс). Начало возбуждения происходит в синусовом узле. Собственная частота возбуждения синусного узла составляет около 75 ударов в минуту, но она регулируется воздействием а) симпатической и парасимпатической нервных систем и б) содержанием углекислоты в плазме крови (и других веществ), в) венозным давлением возврата крови. Концентрация углекислоты и венозное давление повышаются при физической работе мышц. Это регулирование происходит в широких пределах: от полной остановки сердца до 220 ударов в минуту. Типовая ЭКГ представлена на рис 4.9.
В миокарде фронт деполяризации распространяется в виде единого ДЗС поперек мышечных волокон: от эндокарда к эпикарду. Фронт возникает почти сразу по всей внутренней поверхности эндокарда (неодновременность порядка 10 мс.). При этом формируется ДЗС сложной формы, повторяющей форму желудочков сердца. Согласно теореме о потенциале ДЗС наблюдаем мы только сигнал, определяемый замыкающим контуром ДЗС, форма которого определяется геометрией сердца и лишь косвенно ходом процесса деполяризации. По этой причине многие подробности возбуждения миокарда оказываются скрытыми и при расшифровке кардиограмм требуется опыт и искусство врача. Подробнее сигнал ЭКГ рассмотрен в главе 5.2 Сигналы энцефалограммы
Т.к. в формировании ЭЭГ принимает участие множество нейронных структур, то наблюдается суммарный сигнал шумового вида. Он периодический и почти синусоидальный, что говорит о кольцеобразном движении возбуждения. Большую роль в наблюдении ЭЭГ имеет метод вызванных потенциалов. По этому методу реакция ЦНС вызывается внешними стимулами и наблюдаемый сигнал имеет характер синхронизированных откликов. Накопление 100 – 1000 откликов позволяет их выделить в шумах. Вид откликов характеризуется многими пиками, каждый из которых отображает прохождение возбуждения отдельной структурой ЦНС. (рис 4.9 б,в,г)
|
|||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-06-14; просмотров: 127; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.5 (0.011 с.) |
|||||||||||||||||||||||||||||||

 Более легкие ионы проникают сквозь мембрану, а более тяжелые не проникают. Возникает поляризация мембран c потенциалом 60-80 мВ. В реальности процессы более сложные, т.к. присутствуют ионы многих типов и не очень понятны действия калий – натриевых насосов.
Более легкие ионы проникают сквозь мембрану, а более тяжелые не проникают. Возникает поляризация мембран c потенциалом 60-80 мВ. В реальности процессы более сложные, т.к. присутствуют ионы многих типов и не очень понятны действия калий – натриевых насосов.
 Рассмотрим особенности формы импульса при деполяризации сферической клетки для униполярного внешнего электрод А, как показано на рис 4.6. На рисунке представлена структура клетки нерва. Возбуждение от одного из сенсорных дендритов достигает клетки и начинается ее деполяризация. Процесс можно представить как "раскрывающуюся" сферическую клетку от точки начального возбуждения. Согласно рассмотренной выше теореме о потенциале ДЗС электрод А будет наблюдать начало импульса «раскрытия» пропорционально увеличивающемуся пространственному углу W, опирающемуся на контур L деполяризованного участка клетки. Положим, что радиус этого контура нарастает линейно во времени t. Тогда угол W будет изменяться по параболическому закону кt2 (к - константа). Такая форма будет сохраняться, пока не достигается максимум. Затем импульс перейдет в характерную для разряда конденсатора экспоненциальную форму. Нам примерно известна удельная емкость мембраны клетки Суд=(1-10) мкФ/см2. Тогда мембранная емкость половины клетки диаметром D будет равна S*Cуд=0.5ПD2Cуд. Разряд емкости происходит на сопротивление внешнего к клетке пространства, его величина примерно равна R=r/4πD (см. раздел 2. Монополь). Постоянная времени разряда t будет t = Суд*D*r/2 (r=1000 Ом*метр, D=10-4м). Для принятых значениях t =0,125-1.25 мс. Эти величины хорошо регистрируются, однако амплитуды импульсов незначительны для реальных расстояний до электродов (единицы мкВ).
Рассмотрим особенности формы импульса при деполяризации сферической клетки для униполярного внешнего электрод А, как показано на рис 4.6. На рисунке представлена структура клетки нерва. Возбуждение от одного из сенсорных дендритов достигает клетки и начинается ее деполяризация. Процесс можно представить как "раскрывающуюся" сферическую клетку от точки начального возбуждения. Согласно рассмотренной выше теореме о потенциале ДЗС электрод А будет наблюдать начало импульса «раскрытия» пропорционально увеличивающемуся пространственному углу W, опирающемуся на контур L деполяризованного участка клетки. Положим, что радиус этого контура нарастает линейно во времени t. Тогда угол W будет изменяться по параболическому закону кt2 (к - константа). Такая форма будет сохраняться, пока не достигается максимум. Затем импульс перейдет в характерную для разряда конденсатора экспоненциальную форму. Нам примерно известна удельная емкость мембраны клетки Суд=(1-10) мкФ/см2. Тогда мембранная емкость половины клетки диаметром D будет равна S*Cуд=0.5ПD2Cуд. Разряд емкости происходит на сопротивление внешнего к клетке пространства, его величина примерно равна R=r/4πD (см. раздел 2. Монополь). Постоянная времени разряда t будет t = Суд*D*r/2 (r=1000 Ом*метр, D=10-4м). Для принятых значениях t =0,125-1.25 мс. Эти величины хорошо регистрируются, однако амплитуды импульсов незначительны для реальных расстояний до электродов (единицы мкВ). Диаметр нашего ДЗС примерно равен диаметру нейрона, т.е. не более 0.1мм. На реальных расстояниях от приемных электродов этот ДЗС с высокой точностью эквивалентен диполю, момент диполя направлен по оси нейрона, с плюсовым напряжением по направлению движения фронта. Зная скорость и расстояние от нейрона до приемного электрода, легко определить наблюдаемую форму импульса. На рис 4.7 показаны реальные формы сигналов нейронов. Наблюдаемую амплитуду можно оценить по формуле раздела 2 Диполь: U( R,φ ) = = V (D*ℓ /2R 2)*cos φ, где V =80мВ, D =0.1мм, ℓ= 20*10-3 мм, R- расстояние до активного электрода от бегущего ДЗС. R и cosφ необходимо выразить через общий параметр t.
Диаметр нашего ДЗС примерно равен диаметру нейрона, т.е. не более 0.1мм. На реальных расстояниях от приемных электродов этот ДЗС с высокой точностью эквивалентен диполю, момент диполя направлен по оси нейрона, с плюсовым напряжением по направлению движения фронта. Зная скорость и расстояние от нейрона до приемного электрода, легко определить наблюдаемую форму импульса. На рис 4.7 показаны реальные формы сигналов нейронов. Наблюдаемую амплитуду можно оценить по формуле раздела 2 Диполь: U( R,φ ) = = V (D*ℓ /2R 2)*cos φ, где V =80мВ, D =0.1мм, ℓ= 20*10-3 мм, R- расстояние до активного электрода от бегущего ДЗС. R и cosφ необходимо выразить через общий параметр t.