
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Динамика вср специалистов экстремального профиля при тренировке в газово-дымовой камереСодержание книги
Поиск на нашем сайте
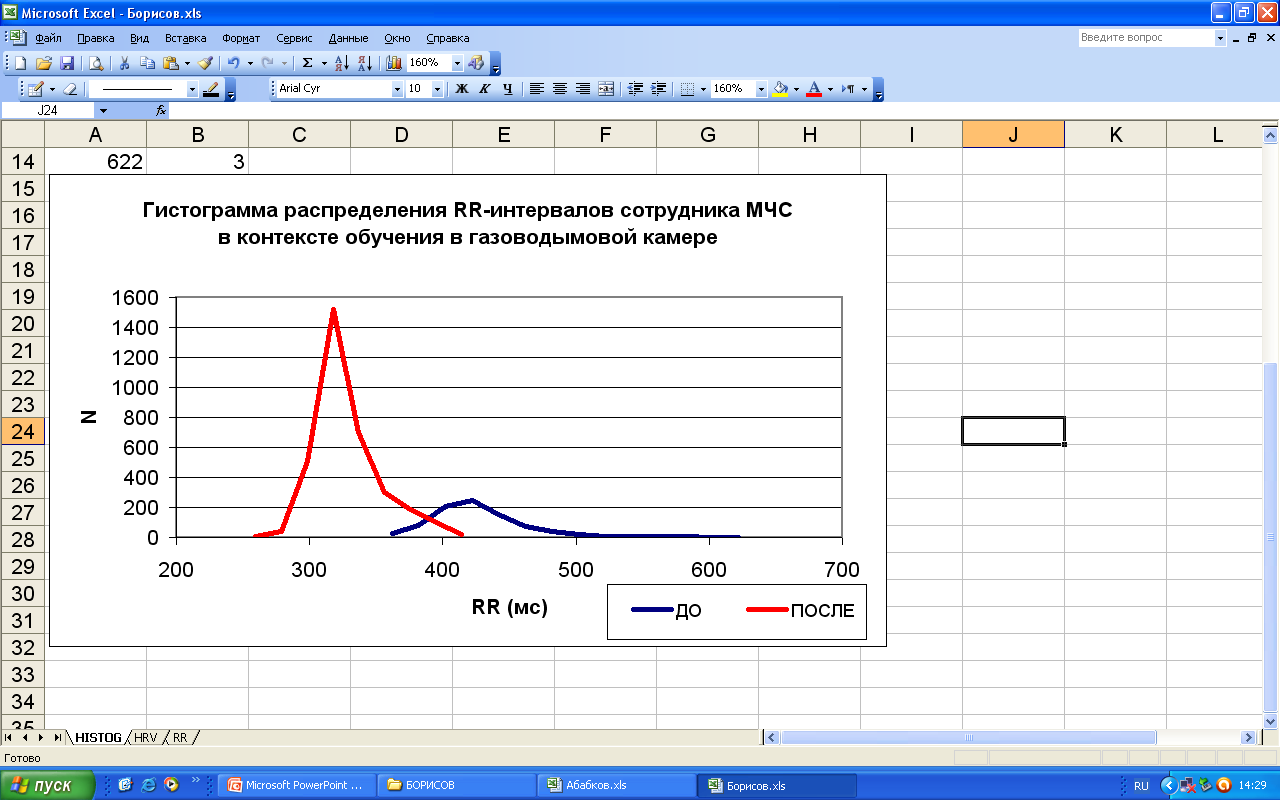
В данной серии экспериментов проводился мониторинг сердечного ритма сотрудников экстремального профиля в процессе тренировки в газово-дымовой камере в контексте естественной деятельности. Тренировка в газово-дымовой камере включает в себя физическую нагрузку высокой интенсивности в условиях ограниченных ресурсов (недостаток кислорода, сниженная видимость, ограничения времени и др.), поэтому является моделью чрезмерных физических нагрузок – стресса. Выборку составили 72 человека мужского пола в возрасте от 19 до 42 лет. На рисунке 66 представлен пример динамики RR-интервалов при входе испытуемого в газово-дымовую камеру (красным маркером отмечен момент входа испытуемого в газово-дымовую камеру). Важно отметить, что в 100% случаев у испытуемых наблюдается значимое снижение RR-интервалов после входа в газово-дымовую камеру, при этом в 63% случаев зафиксированы случаи аритмии и экстрасистолии.
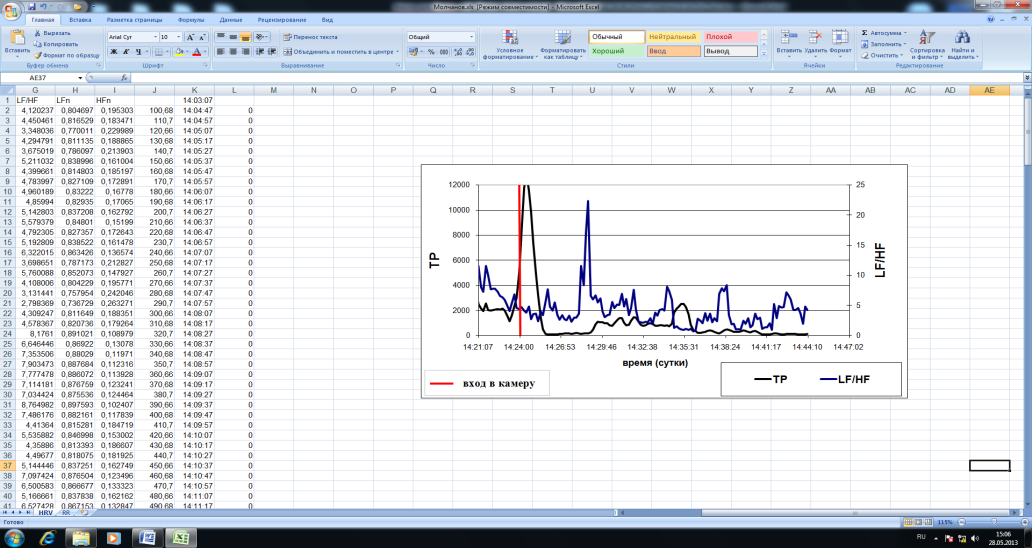
Рис. 66. Пример динамики RR-интервалов в контекстах до входа и внутри газово-дымовой камеры. При анализе динамики спектральных показателей ВСР у испытуемых была выявлена характерная динамическая структура общей мощности спектра ВСР и ИВБ (97% случаев), а именно снижение общей мощности спектра ВСР и возрастание ИВБ (ТР↓,LF/HF↑) при начале тренировки. Данная динамическая структура следует за всплеском общей мощности колебаний, который согласован по времени с моментом входа в камеру (Рисунок 67).
Рис. 67. Средняя динамика спектральных показателей ВСР испытуемых при входе в газово-дымовую камеру (ось х – масштабированное время в % относительно времени пребывания в газово-дымовой камере, принятого за 100%). Важно отметить, что не усредненная – индивидуальная динамика общей мощности спектра ВСР и ИВБ более нестационарна. Внутри газово-дымовой камеры часто наблюдаются дополнительные относительно периодические модуляции ИВБ, нередко встречаются повторные крестообразные структуры ТР↓,LF/HF↑ (Рисунок 68). По всей видимости, это может быть связано с факторами внешнего контекста внутри камеры.
Рис. 68. Пример динамики общей мощности спектра ВСР и ИВБ испытуемого в контексте тренировки в газово-дымовой камеры (красным маркером отмечен момент входа в камеру, зеленой заливкой выделена повторная динамическая структура ТР↓,LF/HF↑).
Таким образом, стресс-реакция, связанная с чрезмерной физической нагрузкой, сопровождается 2-стадийной динамикой спектральных показателей ВСР: 1. возрастание общей мощности спектра ВСР и ИВБ, 2. снижение общей мощности спектра ВСР на фоне возрастания ИВБ. При этом напряжение регуляторных систем, связанное с физической нагрузкой, сопровождается согласованными по времени возрастанием общей мощности спектра ВСР и ИВБ и последующим их снижением. Обсуждение результатов
Результаты исследования динамики вегетативной регуляции кардиоритма при нагрузках разного рода, в первую очередь, воспроизводят и подтверждают данные о высокой чувствительности системы регуляции к различным изменениям во внутреннем и внешнем контекстах живой системы [279]. Использование нового инструментария – системы беспроводной регистрации сердечного ритма, который был разработан специально под задачи измерения режимов работы живой системы в условиях естественной деятельности, открыло новые возможности для исследований [291; 293]. В результате многообразных экспериментальных серий измерений в различных контекстах естественной деятельности нам удалось выяснить, что динамика параметров вегетативной регуляции кардиоритма является нелинейной, квазипериодической и не имеет прямых взаимосвязей с режимом вегетативной регуляции в стационарных контекстах – сидя, лежа. Это ставит под сомнение адекватность такой характеристики, как вегетативный статус человека, который во многих работах трактуется как режим вегетативной регуляции, типичный для данного субъекта [305; 237]. Мы считаем, что все режимы вегетативный регуляции специализированы под конкретные функциональные контексты. Конечно, нет оснований полностью отрицать возможность выявления зависимостей между показателями вегетативной регуляции в покое и в контексте естественной деятельности. Однако их, видимо, стоит искать персонально, для конкретного человека посредством многократного длительного мониторинга, что, возможно, позволит с определенной точностью предсказывать вегетативные реакции конкретного человека в богатой стимульной среде естественной деятельности.
Использование алгоритмов спектрального анализа, специализированных для неравномерных временных рядов (каким, собственно и является последовательность RR-интервалов), позволило выявить новый, не рассматриваемый в классической физиологии и медицине, диапазон спектра ВСР – очень высокочастотные колебания от 0,6 до 2 Гц. Показатели данного диапазона оказались чувствительными к функциональному состоянию стресса и уровнем активности ЭОС. Поэтому есть все основания полагать, что в структуре нервно-гуморальной регуляции сердечного ритма присутствуют механизмы быстрой, краткосрочной регуляции. Математические методы динамического спектрального анализа позволяют масштабировать дискретность получаемых параметров, поэтому появляется возможность наблюдать изменения структуры нервно-гуморальной регуляции сердечного ритма, согласованные по времени с динамикой внешних событий. Это шаг к исходному пониманию состояний как функции от времени, который приближает к возможности выявлять закономерные динамические структуры и предсказывать динамику событий эндогенного контекста живой системы в более короткие сроки. В нашей работе также продемонстрировано подтверждение того, что для спектрального анализа ритмограмм необходимо применять методы, специализированные под неравномерно дискретизированные сигналы. Только в таком случае, без применения алгоритмов интерполяции, в получаемых спектрах ритмограмм наблюдается очень высокочастотный компонент (0,6-2 Гц), характеристики которого, как оказалось, чувствительны для распознавания функциональных состояний и наркозависимости. Этот результат дает возможность использовать для анализа более короткие фрагменты ритмограмм (15-20 с), чем приняты в классических методах анализа ВСР (300 с). Полученные нами результаты сравнения динамики вегетативной регуляции кардиоритма при когнитивной нагрузке подтверждают литературные данные, в которых демонстрируются вегетативные эффекты когнитивного напряжения, отражающиеся в снижение вариабельности сердечного ритма [105; 106; 26; 131; 151; 173; 132; 65; 260; 12; 103; 202; 211; 1; 9; 157; 27; 84]. Принципиально новым в нашем исследовании стало сравнение разных контекстов когнитивной нагрузки, которые отличаются между собой не степенью сложности (все контексты задействовали первичные когнитивные функции), а характером целевой задачи. Соответственно, такой подход дал очень интересный результат, который заключается в том, что режим вегетативной регуляции меняется и при смене типа когнитивной задачи. Это указывает на наличие очень тонких механизмов регуляции. Кроме того, показаны быстрые изменения сердечного ритма, согласованные с динамикой информационного образа и движением живой системы в когнитивном пространстве по циклу определенность-неопределенность. Увеличение уровня дезорганизации сердечного ритма в контексте неопределенности, возможно, связано с активизацией дополнительных нейрональных модулей обработки информации и их проекциями на систему нервно-гуморальной регуляции сердечного ритма. Результаты, полученные нами при исследовании связей между вегетативной регуляцией кардиоритма и эмоциональным состоянием, также находят подтверждение в литературных данных. Угнетенное эмоциональное состояние (в нашем случае повышение уровня эмоциональной дезадаптации), нарушения эмоционально-волевой сферы сопровождаются редукцией вегетативной регуляции кардиоритма [10; 37; 207; 128]. Относительно этого факта возникает вопрос первичности, так как исследования вегетативных расстройств (вегетососудистая дистония) демонстрируют ухудшение эмоционального состояния больных [252] (как будто при других болезнях наблюдается улучшение эмоционального состояния). По-видимому, в основе механизмов этих эффектов лежит взаимное влияние, и даже включенность в единый системный процесс двух модулей – эмоционального и вегетативного.
Вообще, снижение вариабельности сердечного ритма как признак наличия какой-либо болезни встречается во многих работах [58; 43; 14]. Список болезней при этом очень широкий: нарушения сердечно-сосудистой системы (дисфункция миокарда, тетраплегия, гипертензия, застойная недостаточность кровообращения, хроническая митральная регургитация, кардиомиопатия, желудочковые аритмии, наджелудочковые аритмии и др.), психоневрогологические нарушения (посттравматическое стрессовое расстройство, депрессия, тревожность), онкологические заболевания, инфекционные заболевания (грипп, ОРВИ). Важным является то, что в исследованиях экспериментальную выборку составляют больные с тем или иным диагнозом, а контрольную – здоровые испытуемые, то есть, нет попыток дифференцировать по вариабельности сердечного ритма больных с разными диагнозами. По всей видимости, снижение вариабельности сердечного ритма является неспецифическим признаком наличия болезни. Тогда, предполагается возможным расширить знаменитую триаду Г. Селье – комплекс симптомов, отражающих стресс: гипертрофия надпочечников, инволюция тимуса, язвообразование в ЖКТ, - включив в нее четвертым пунктом снижение вариабельности сердечного ритма. Наши данные позволяют предположить, что не только автономный контур регуляции снижает активность при повышении уровня эмоциональной дезадаптации, но и центральный контур меняет свою структуру: циклы регуляции становятся более локализованными. Можно предположить, что это связано с формированием узкой доминанты активности нейрональных структур, задействованных в эмоциональных процессах и процессах вегетативного обеспечения поведения. Полученные нами данные о динамике вегетативной регуляции кардиоритма до и после физической нагрузки воспроизводят результаты многих предыдущих исследований, которые демонстрируют снижение уровня вариабельности сердечного ритма спортсменов после тренировки или после соревновательного периода [31; 146; 192; 61; 205]. Принципиально новыми стали результаты непрерывного мониторинга сердечного ритма спортсменов в процессе тренировки, синхронизированные по времени с динамикой тренировочной нагрузки. При их анализе было выявлено, что максимальное напряжение наблюдается на этапе короткого по времени перехода от низкоинтенсивной активности к высокоинтенсивной. По всей видимости, это напряжение регуляторных систем связано с необходимостью быстрой актуализации большого количества ресурсов. Наличие в структуре тренировочного процесса частых переходных моментов (от низкоинтенсивной к высокоинтенсивной нагрузке и наоборот) ведет к срыву механизмов нейрогуморальной регуляции сердечного ритма и проявляется в наличии случаев аритмии и экстрасистолии.
Важный практически ценный результат был получен при попытке выявить структуру связей между исходным состоянием и эффективностью деятельности спортсменов. Отсутствие среднестатистических, универсальных и наличие персонифицированных зависимостей, вероятно, объясняется тем, что совокупность функциональных систем, реализующая эффективное выполнение целевой задачи, не одинакова у спортсменов. Это значит, что и значения показателей, отражающих оптимизированное ресурсообеспечение сформированной иерархии функциональных систем, будут разные. То есть структура функциональных модулей, включенных в систему, оптимизированную для выполнения профессиональных задач, персональная, а не универсальная и сформирована в процессе индивидуального опыта обучения и тренировок. Таким образом, предсказание эффективности в игровой деятельности по психофизиологическим показателям должно производиться на основе индивидуальных моделей оптимальной реализации профессиональных навыков. Соответственно, выбор необходимых мер для активизации нужного игрока и приведения его в индивидуальную зону максимальной эффективности по параметрам функционального состояния должен осуществляться на основе индивидуальных психофизиологических профилей, соответствующих максимальной эффективности. На основании этих результатов можно предположить, что они воспроизведутся и в других областях деятельности, не обязательно связанных с моторной активностью. Такой подход (персонифицированный мониторинг в контексте естественной деятельности) открывает новые возможности для оптимизации условий и режимов учебной и профессиональной деятельности конкретно для каждого человека. Таким образом, при предъявлении разнообразных когнитивных, моторных и эмоциональных нагрузок, не выходящих на уровень стрессогенности, мы получили различные варианты динамики ВСР, общим для которых стало синхронное нарастание ИВБ и общей мощности спектра ВСР. При поиске динамического вегетативного маркера стресса в работе исследована динамика вегетативной регуляции кардиоритма в широкой группе контекстов естественной деятельности, которые содержат разнообразные стресс-факторы: информационные, эмоциональные, физические. В итоговой таблице 17 представлены обобщенные результаты наших экспериментов. По этим данным можно заключить наличие динамической структуры общей мощности спектра ВСР и ИВБ, которая возникает в ситуациях чрезмерного рассогласования между целевой функцией и наличествующим результатом независимо от природы контекста (эмоциональный, информационный, моторной активности). Это дает основания утверждать, что данный паттерн активности систем регуляции сердечного ритма является маркером стресс-реакции.
Таблица 17. Последовательная динамика показателей ВСР в контекстах естественной деятельности («á»-возрастание; «â»-снижение; «-» - не меняется).
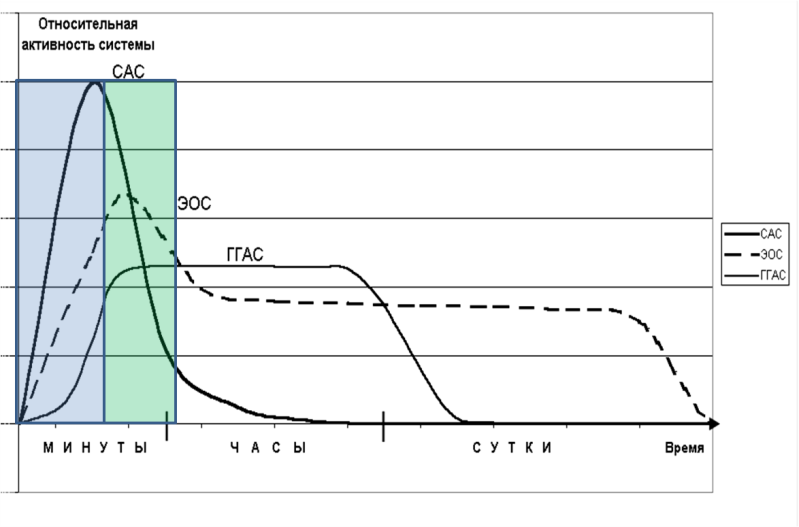
Напомним, что эта структура включает следующие последовательные фазы: 1) возрастание обще мощности спектра ВСР и ИВБ, 2) возрастание ИВБ при снижении общей мощности спектра ВСР (Рисунок 69, А). А Рис. 69. Обобщенная схема динамики спектральных показателей ВСР: общей мощности спектра и ИВБ при стресс-активации (А) и физической нагрузке (Б). Важно, что по динамике показателей общей мощности спектра ВСР и ИВБ можно разделить стресс-активацию от напряжения регуляторных систем, связанного с моторной активностью. Различие заключается в том, что моторная активность в контексте спортивной тренировки сопровождается однонаправленными изменениями этих показателей. Стоит отметить, что изменения длительности RR-интервалов и значений ЧСС в этом случае неинформативны: и при стрессе и при моторной активности наблюдается их уменьшение. Естественно, встает вопрос о психофизиологических механизмах такой динамики показателей ВСР при стрессе. Чтобы попробовать в них разобраться, для начала проанализируем, от чего зависит показатель мощности колебаний RR-интервалов. Во-первых, имеет значение диапазон изменений RR-интервалов, который численно можно выразить в формуле RRmax-RRmin. По сути, этот показатель отражает количество степеней свободы в сигнале. Соответственно, чем он выше, тем больше возможных регуляторных команд могут влиять на сигнал и отражаться в нем различными амплитудно-фазовыми модуляциями, которые дают прирост мощности в спектре. Если данный диапазон узкий, что наблюдается при высоких значениях ЧСС, то команды могут поступать, но не отрабатывать. Во-вторых, колебания сердечного ритма содержат модуляции, связанные с командами от системы нервно-гуморальной регуляции. Соответственно, мощность спектра в таком сигнале может зависеть от сложности структуры управляющей системы. Снижение мощности, в таком случае, связано либо со снижением активности центров управления (стало меньше команд), или с реструктуризацией системы управления в сторону локализации источника команд (команд столько же, но они исходят только от одного модуля системы, при этом прочие модуляции, отражающиеся в других частотных диапазонах спектра, подавляются). Нельзя исключать из рассмотрения и совместное действие обоих алгоритмов. Таким образом, можно объяснить первую фазу динамики общей мощности спектра ВСР и ИВБ в стресс-активации и моторной активности (TP↑LF/HF↑) активизацией всех контуров системы нервно-гуморальной регуляции сердца. Это, в свою очередь, может быть обусловлено потребностью в короткие сроки активировать максимум ресурсов и перестроить систему в новый режим (физической нагрузки). Тогда вторая фаза динамики общей мощности спектра ВСР и ИВБ при стрессе (ТР↓LF/HF↑) соответствует переходу системы управления в редуцированный режим, структура системы упрощается. По сути, это согласовано с представлениями о стрессе, как о древней, сугубо упрощенной реакции [280; 296]. Как показано в исследованиях [287], при стрессе стираются индивидуальные различия. Снижение же общей мощности спектра ВСР и ИВБ при моторной нагрузке, по-видимому, связано с сильным ограничением степеней свободы в регуляции работы сердца. Перейдя в режим максимального ресурсообеспечения мышечного аппарата организма, центры регуляции становятся не способными к дополнительным модуляциям процесса. Обобщенно данные механизмы представлены в схеме на рисунке 70. А. В. Рис. 70. Система управления сердечным ритмом в покое (А), при стрессе (Б), при моторной активности (В). Стрелки отображают богатство нисходящих и восходящих нейровисцеральных связей. Переводя приведенную логику на язык теории функциональных систем, можно сказать, что мощность спектра ВСР отражает сложность структуры иерархии функциональных систем, задействованных в данный момент. Тогда снижение общей мощности спектра ВСР при стрессе ещё раз подтверждает предположение о том, что стресс является низкодифференцированной эволюционно древней функциональной системой. Опираясь на 3-компонентную теорию стресса [280] и данные эмпирических исследований вегетативной регуляции кардиоритма при нейрохимических и гуморальных изменениях организма [263; 212; 119; 45], мы предполагаем, что первая фаза динамики связана с активацией всех регуляторных контуров, и особенно симпатической подсистемы, и согласована с первой стадией стресса – активацией САС. «Симпатоадреналовая система – это нейроэндокринный комплекс, направленный на активацию висцеральных систем» [цит. по: 285]. Симпатический компонент автономной нервной системы (основные медиаторы – норадреналин и адреналин) и гормоны мозгового вещества надпочечников (адреналин и норадреналин) обеспечивают комплекс одноплановых реакций: активизацию сердечной деятельности и перераспределение кровотока «в пользу» скелетной мускулатуры, сердца и мозга. Вторая фаза динамики может быть связана с включением быстрого компонента динамики ЭОС ещё на первой стадии стресса. «Эндогенная опиоидная система - это комплекс морфиноподобных пептидов (эндорфины, энкефалины, динорфины и др.) с выраженным тормозным действием на большинство систем организма, реализующимся через опиатные рецепторы (в основном, через пресинаптическое торможение)» [цит. по: 285] (Рисунок 71). Дополнительным аргументом в пользу такой трактовки динамики являются характерные для нее времена (от 1 до 20 мин.).
Рис. 71. Соответствие фаз динамической структуры общей мощности спектра ВСР и ИВБ (Б) при стрессе и первой стадии динамики стресс регуляторных систем (А) [по: 280; 244]. Безусловно, полученный в ходе исследования динамический маркер стресса необходимо воспроизвести в лабораторных экспериментах на животных с использованием методов фармако-физиологического анализа. Важно обратить внимание на противоречие, которое может возникнуть при трактовке наших результатов и в принципе наблюдается в кругу исследователей данной области [82]. Противоречие обнаруживается при сопоставлении двух признаков активности симпатического отдела ВНС – мощности низкочастотного компонента спектра ВСР (LF) и ЧСС. Так, принято считать, что повышение ЧСС и увеличение показателя LF отражают активность симпатической ВНС. В нашем случае при повышении ЧСС наблюдается снижение мощности LF компонента (например, у спортсменов), то есть два признака противоречат друг другу. Мы нашли следующее объяснение такого эффекта. Эксперименты в исследованиях LF компонента, как признака, отражающего активность симпатической ВНС, исторически проводились в контекстах покоя или на наркотизированных животных, то есть не рассматривались активные контексты. В таких случаях действительно наблюдаются прямые корреляции между мощностью LF и ЧСС, которые исчезают при рассмотрении активных контекстов. Однако, в контексте покоя можно наблюдать периодическое повышение ЧСС при высокой мощности HF компонента спектра ВСР, который отражает активность парасимпатического звена ВНС (например, при выраженной дыхательной волне в ритмограмме). В итоге получается, что изменения ЧСС и мощности LF не всегда связаны именно с активностью симпатического отдела ВНС. По всей видимости, они отражают разные режимы симпатического воздействия на сердечный ритм. Тогда LF отражает фазические модуляции сердечного ритма симпатической ВНС в контексте покоя, а повышение ЧСС – однократное сильное возрастание симпатической активации в активном контексте. То есть, отвечая на вопрос: является ли повышение ЧСС маркером симпатической активации, следует уточнить: насколько и за какое время снизилось ЧСС, а в отношении LF – компонента при трактовке результатов следует учитывать характеристики контекста, в котором проводились измерения. По сути, такие противоречия являются лишь малой частью крупной проблемы выработки норм по физиологическим параметрам и границ их применения. Завершая обсуждение, перейдем к роли ЭОС в регуляции сердечного ритма. Известно, что длительное употребление опиатных наркотиков ведет к разрушению рецепторного аппарата ЭОС [190]. Поэтому наркозависимые больные в период отмены являются испытуемыми, у которых нарушена регуляторная функция ЭОС. Сравнивая их показатели с показателями здоровых испытуемых, мы выделили характерные особенности (Рисунок 72). В частности, принципиально важным оказалось сниженение эффективности нервно-гуморальной регуляции сердечного ритма у наркозависимых. Мощностные характеристики ВСР в экспериментальной группе значимо ниже во всех контекстах (покой, когнитивная нагрузка). Такой эффект может иметь два объяснения. Во-первых, из структуры нервно-гуморальной регуляции сердечного ритма исключается одна из базовых нейрохимических систем мозга – ЭОС. Тогда система управления сердечным ритмом существенно сужается. Во-вторых, нарушение ЭОС ведет к устранению тормозных механизмов, ввиду чего симпатоадреналовые воздействия становятся гиперактивированными. Такой эффект также может быть связан и с изменениями на информационном уровне живой системы. Исходя из того, что ЭОС является одним из главных звеньев в управлении сигналами о рассогласовании, а конкретнее, ингибитором таких сигналов, можно предположить, что нарушение ЭОС ведет к возникновению многочисленных сигналов о рассогласовании, что в свою очередь приводит к снижению степеней свободы в настройках режима вегетативного обеспечения организма. А. В. Рис. 72. Динамика параметров ВСР (А – TP (мс2); Б – LF (мс2); В – HF (мс2)) в 6 пробах (ось х – контекст: 0. – лежа; 1– компьютерная латерометрия; 2 - компьютерная кампиметрия; 3 - тест Струпа; 4 - тест «Часы»; 5 – тест «Управление углом наклона отрезка»; 6 - измерение времени сенсомоторной реакции) в группах: 0-здоровые (синим), 1- наркозависимые (красным). Результаты сравнения показателей когнитивных функций наркозависимых и здоровых испытуемых, по сути, подтверждают такую трактовку. Так, в тесте «Часы» наркозависимые в большинстве случаев переходят к следующему заданию только при достижении нулевой ошибки «установленного времени», то есть при достижении ситуации отсутствия объективного рассогласования между заданным и установленным временем, чего не наблюдается у здоровых испытуемых. Это ведет к выводу о том, что у наркозависимых испытуемых в сравнении со здоровыми ниже порог допустимого рассогласования, ввиду чего одни и те же объективные рассогласования субъективно воспринимаются по-разному. Такое утверждение подтверждают результаты измерения времени простых сенсомоторных реакций. Мы выявили, что в группе наркозависимых испытуемых в сравнении со здоровыми значимо выше время моторного компонента реакции, то есть, проще говоря, они дольше удерживают клавишу, реагируя на звуковой стимул. Если учесть, что выполнение поведенческого акта заканчивается при устранении сигнала о рассогласовании, то наблюдаемая разница времен может быть объяснена аналогично нарушением ЭОС – сигналы о рассогласовании дольше не устраняются. Такой же эффект воспроизводится и при сравнении времени сенсомоторной реакции испытуемых в тесте Струпа, в котором реализована модель когнитивного конфликта (в пробах, в которых значение слова не совпадает с цветом букв). Аналогично, рассогласование между смыслом слова и цветом букв субъективно для наркозависимых является более выраженным и тормозит процесс принятия решения при выборе.
Заключение
Главной задачей исследования был анализ непрерывной динамики вегетативной регуляции кардиоритма при разнородных нагрузках в контексте естественной деятельности и выявление динамических структур показателей вегетативной регуляции, специфичных для стресс-реакции – маркеров стресса. В результате удалось выяснить, что снижение общей мощности спектра ВСР на фоне возрастания индекса вегетативного баланса, которое следует за их совместным возрастанием, сопровождает системные процессы в ответ на сигнал о чрезмерном рассогласовании в контекстах эмоциональной, информационной и физической нагрузки. Данная динамическая структура показателей ВСР отвечает всем выделенным критериям к маркерам стресса: 1. динамичностью, 2. неспецифичностью к природе стресс-фактора, 3. стадийностью. В результате в нашей работе продемонстрированы прямые доказательства того, что функциональная система стресса на психофизиологическом обладает свойствами: 1. неспецифичности. Одна и та же структура режима вегетативной регуляции воспроизводится при эмоциональных (контекст публичного выступления), информационных (внезапные маневры при управлении автотранспортом, ошибки в компьютерных играх и др.) и физических (контекст тренировки в газово-дымовой камере) стресс-факторах. 2. редуцированности. Общая мощность спектра вариабельности сердечного ритма и мощности в низкочастотном и высокочастотном диапазонах спектра снижаются в разы. 3. динамичности. Стресс-реакция и ее отражение в вегетативной регуляции сердечного ритма имеют закономерную последовательность смены режимов работы живой системы. Вторым важным аспектом работы стали измерения в контексте естественной деятельности. Разработанная для этого система беспроводной регистрации кардиоритма на основе миниатюрных сенсорных платформ дала возможность получать информацию о динамических режимах работы живой системы без нарушения и искажения формируемых функциональных систем дополнительными компонентами (лабораторные условия, наличие экспериментатора и др.). Это еще один шаг на пути отступления от парадигмы реактивности, в которой рассматриваются реакции исследуемого объекта на предъявляемые, сформированные экспериментатором, стимулы, к парадигме активности, в которой анализируются автономные, самостоятельно конструируемые поведенческие акты и их физиологическое обеспечение [229]. В частности, в нашей работе показано, что спектр режимов живой биологической системы в контексте естественной деятельности значительно шире, чем таковой в стационарных контекстах, формируемых в лабораторных условиях. Выводы
1. Технология регистрации кардиоритма на основе беспроводных сенсорных сетей обеспечивает согласованное измерение экзогенного и эндогенного контекстов при когнитивных, эмоциональных и физических нагрузках с минимизацией рисков нарушения целостности функциональных систем человека. 2. У здоровых людей, находящихся в режиме повседневной активности, вегетативным отображением эмоциональной дезадаптации является снижение активности центрального и автономного контуров регуляции сердечного ритма. Отсутствие эмоциональной дезадаптации в течение суток поддерживается динамическими перестройками режима вегетативной регуляции. 3. В контексте когнитивной нагрузки вегетативным отображением уровня неопределенности информационного образа является уровень дезорганизации сердечного ритма. Адаптационные процессы в контексте оптимальных когнитивных нагрузок отображаются в динамических изменениях режима вегетативной регуляции кардиоритма при смене целевой функции. 4. Адаптационные процессы при интенсивной физической нагрузке (не экстремальной) отображаются в согласованном снижении общей мощности спектра вариабельности сердечного ритма и индекса вегетативного баланса. 5. Вегетативные отображения адаптационных и дезадаптационных процессов отличаются по степени изменчивости динамических режимов кардиоритма: • адаптационные процессы отображаются в многообразии режимов вегетативной регуляции, связанными со спецификой нагрузки, • дезадаптационные процессы отображаются в специфическом редуцированном режиме вегетативной регуляции, который инвариантен к природе экстремальной нагрузки, ситуативным и индивидуальным особенностям. 6. Вегетативным отображением дезадаптационных процессов при экстремальных когнитивных, эмоциональных и физических нагрузках является снижение общей мощности спектра вариабельности сердечного ритма, согласованное по времени с возрастанием индекса вегетативного баланса. 7. У наркозависимых испытуемых отсутствуют изменения режима вегетативной регуляции при смене целевой функции в контексте когнитивной нагрузки. Таким образом, снижение активности эндогенной опиоидной системы ведет к нарушению адаптационных процессов при оптимальных когнитивных нагрузках. Список сокращений
Список литературы 1. Aasman, J., Wijers, A.A., Mulder, G., Mulder, L.J. Measuring mental fatigue in normal daily working routines / In: P.A. Hancock, N. Meshkati (Ed.) Human mental workload - Amsterdam: North-Holland, 1988. - P. 117-138. 2. Ahs, F., Sollers, J.J., Furmark, T., Fredrikson, M., Thayer, J.F. High-frequency heart rate variability and corticostriatal activity in men and women with social phobia // Neuroimage. - 2009. – N 47. – P. 815–820. 3. Akil, H., Watson, S.J. The role of endogenous opiates in pain control // Life Sci. Res. Repts. - 1980. - № 17. - P. 201-222. 4. Akinola, M., Mendes, W.B. The dark side of creativity: biological vulnerability and negative emotions lead to greater artistic creativity // Pers. Soc. Psychol. Bull. - 2008. – N 34. – P. 1677–1686. 5. Akselrod, S. Components of heart rate variability // Heart rate variability. - N.Y.: Armonk.,1995. - P. 146-164. 6. Akselrod, S., Gordon, D., Ubel, F.A. Power spectrum analysis of heart rate fluctuation: A quantitative probe of beat-to-beat cardiovascular control // Science. - 1981. - Vol. 213, N 4503. - P. 220-222. 7. Allen, A.P., Kennedy, P.J., Cryan, J.F., Dinan, T.G., Clarke, G. Biological and psychological markers of stress in humans: Focus on the Trier Social Stress Test // Neuroscience and Biobehavioral Reviews. – 2014. - N 38. – P. 94–124. 8. Altemus, M., Redwine, L.S., Leong, Y., Frye, C.A., Porges, S.W., Carter, C.S., Responses to laboratory psychosocial stress in postpartum women // Psychosom. Med. - 2001. – N 63. P. 814–821. 9. Althaus, M., Mulder, L.J., Mulder, G., Van Roon, A.M., Minderaa, R.B. Influence of respiratory activity on the cardiac response pattern to mental effort // Psychophysiology. – 1998. - Vol. 35., N 4. - P. 420-430. 10. Appelhans, B.M., Luecken, L.J. Heart rate variability as an index of regulated emotional responding // Rev. Gen. Psychol. - 2006. – N 10. – P. 229–240. 11. Aysin, B., Aysin, E. Effect of Respiration in Heart Rate Variability (HRV) // Analysis. Conf Proc IEEE Eng. Med. Biol. Soc. – 2006. –N 1(1). – P. 1776-1779. 12. Backs, R.W., Seljos, K.A. Metabolic and cardiorespiratory measures of mental effort: the effects of level of difficulty in a working memory task // Int. J. Psychophysiol. – 1994. - Vol. 16., N 1. – P. 57-68. 13. Badra, L.J., Cooke, W.H., Hoag, J.B., Crossman, A.A., Kuusela, T.A., Tahvanainen, K.U., Eckberg, D.L. Respiratory modulation of human autonomic rhythms // Am. J. Physiol. Heart Circ Physiol. - 2001. - Vol. 280., № 6. – P. 2674-2688. 14. Becker, K., Görlach, I., Frieling, T., Häussinger, D. Characterization and natural course of cardiac autonomic nervous dysfunction in HIV-infected patients // AIDS. – 1997. – Vol. 11(6). – P. 751-757. 15. Belova, M.A., Paton, J.J., Morrison, S.E., Salzman, C.D. Expectation modulates neural responses to pleasant and aversive stimuli in primate amygdale // Neuron. – 2007. - N 55. – P. 970–984. 16. Ben Lamine, S., Calabrese, P., Perrault, H., Dinh, T.P., Eberhard, A., Benchetrit, G. Individual differences in respiratory sinus arrhythmia – Am. J. Physiol. Heart Circ Physiol. – 2
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2019-05-20; просмотров: 169; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.226.163.23 (0.032 с.) |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




 Б
Б 
 Б.
Б. 

 А.
А. Б.
Б. Б.
Б. 




