Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Белки. Аминокислоты -- структурные компоненты белковСтр 1 из 16Следующая ⇒
Биохимия
§ 1. Введение в биохимию § 1.1. БЕЛКИ. АМИНОКИСЛОТЫ -- СТРУКТУРНЫЕ КОМПОНЕНТЫ БЕЛКОВ § 1.2. Строение и классификация аминокислот § 1.3. Уровни структурной организации белковых молекул § 1.4. Физико-химические свойства белков § 1.5. КЛАССИФИКАЦИЯ БЕЛКОВ § 1.6. Углеводы. Классификация углеводов § 1.7. Липиды § 1.8. Витамины § 1.8.1. Жирорастворимые витамины § 1.8.2. Водорастворимые витамины § 1.8.3. ВИТАМИНОПОДОБНЫЕ ВЕЩЕСТВА. § 2. Ферменты § 2.1. Ферменты и неорганические катализаторы § 2.2. Строение ферментов § 2.3. Коферменты § 2.4. Свойства ферментов § 2.5. Номенклатура ферментов § 2.6. Классификация ферментов § 2.7. Механизм действия ферментов § 2.8. Ингибирование ферментативной активности § 3. Обмен углеводов § 3.1. Биологическая роль углеводов § 3.2. Превращение углеводов в пищеварительном тракте § 3.3. Биосинтез и распад гликогена § 3.4. Основные пути катаболизма глюкозы § 3.4.1. Анаэробный гликолиз § 3.4.2. Аэробный гликолиз (гексозодифосфатный путь) § 3.4.3. Гексозомонофосфатный путь § 3.4.4. Глюконеогенез § 4. Обмен липидов § 4.1. Основные липиды организма человека их биологическая роль. § 4.2. Переваривание липидов, ресинтез жира § 4.3. Липопротеины крови § 4.4. Окисление высших жирных кислот § 4.5. Окисление глицерина § 4.6. Биосинтез ВЖК в тканях § 4.7. Обмен холестерина § 5. Обмен белков § 5.1. Переваривание белков § 5.2. Гниение аминокислот, обезвреживание продуктов гниения § 5.3. Метаболизм аминокислот § 5.4. Пути обезвреживания аммиака § 6. Регуляция обмена веществ § 6.1. Сигнальные молекулы § 6.2. Гормоны гипоталамуса § 6.3. ГОРМОНЫ ГИПОФИЗА § 6.4. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ § 6.5. ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ § 6.6. Гормоны половых желез § 6.7. Гормоны надпочечников § 6.8. Гормоны поджелудочной железы § 7. Экзаменационные вопросы
БЕЛКИ. АМИНОКИСЛОТЫ -- СТРУКТУРНЫЕ КОМПОНЕНТЫ БЕЛКОВ БЕЛКИ Белки – это азотсодержащие, высокомолекулярные органические соединения, состоящие из аминокислот, соединенных в цепи с помощью пептидных связей и имеющие сложную структурную организацию. Строение и классификация аминокислот Существуют различные классификации аминокислот, входящих в состав белков в зависимости от признака, положенного в основу их деления на группы. Так по химической природе боковой цепи a-аминокислоты делятся на алифатические (ациклические), ароматические, гетероциклические.
I. Алифатическиеa - аминокислоты 1. Моноаминомонокарбоновые (нейтральные) Алифатические аминокислоты, содержащие в боковой цепи дополнительную функциональную группу. а) гидроксильную группу (оксиаминокислоты) б) карбоксильную группу (моноаминодикарбоновые) в) амидную группу г) аминогруппу (диаминомонокарбоновые) д) серусодержащие II. Ароматические аминокислотыIII. Гетероциклические аминокислоты IV. Иминокислота Вместе с тем в настоящее время общепринятой является классификация аминокислот на основе признаков, свойственных R-группам, в частности, их полярности, т.е. способности R-групп взаимодействовать с водой при биологических значениях рН=7,0. Это обусловлено наличием в боковой цепи аминокислот различных функциональных групп. Так к неполярным гидрофобным группам (водоотталкивающим) относятся алкильные радикалы, бензольное кольцо. Гидрофильными (полярными) группами являются группы: -ОН, -SН, -СООН, - NН2, ядро имидазола. Гидрофильность - это свойство молекул или групп, имеющих полярную природу, легко взаимодействовать с водой. Полярные функциональные группы, способные к ионизации, называют ионогенными, они несут отрицательный заряд, если отщепляют протон или положительный заряд, если его принимают. В связи с этим все 20 аминокислот по свойствам боковой цепи можно разделить на 4 группы: Неполярные гидрофобные, Полярные неионные, В боковой цепи этих аминокислот содержатся неполярные, неионные группы. К данному классу относятся алифатические аминокислоты. Глицин это самая маленькая аминокислота, содержащая в боковой цепи атом водорода, поэтому в белке её остаток не имеет ярко выраженных гидрофобных или гидрофильных свойств. Аминокислоты аланин, валин, лейцин, изолейцин - алифатические неполярные аминокислоты. По мере увеличения боковой цепи увеличивается их гидрофобность. К этой группе относятся также аминокислоты триптофан, фенилаланин, метионин и иминокислота - пролин
Аминокислоты этой группы содержат в боковой цепи неионогенные группы, не способные отдавать или присоединять протон. Боковые группы этих аминокислот растворяются в воде т.к. в их состав входят полярные функциональные группы, образующие водородные связи с водой. К ним относятся серин, треонин, содержащие в боковой цепи спиртовой гидроксил, а также глутамин, аспарагин, содержащие амидные группы. К этой же группе относятся цистеин и тирозин. Эти аминокислоты содержат соответственно тиольную группу и фенольный гидроксил, способные к диссоциации, но при нейтральных значениях рН, поддерживаемых в клетках, эти группы практически не диссоциируют. R- группами. Дополнительную положительно заряженную группу в боковой цепи имеют лизин, аргинин. Кроме того гистидин содержит слабо ионизированную имидазольную группу, поэтому при физиологических колебаниях значений рН (от 6,9 до 7,4) гистидин заряжен либо нейтрально, либо положительно. При увеличении количества протонов в среде имидазольная группа гистидина способна присоединять протон, приобретая положительный заряд, а при увеличении концентрации гидроксильных групп – отдавать протон теряя положительный заряд.
Белковые модули (домены) Обычно белки, образованные одной полипептидной цепью, представляют собой компактное образование, каждая часть которого не может функционировать и существовать отдельно, сохраняя прежнюю структуру. Однако, в некоторых случаях, при большом содержании аминокислотных остатков (более 200), в трехмерной структуре обнаруживается не одна, а несколько независимых компактных областей одной полипептидной цепи. Эти фрагменты полипептидной цепи, сходные по свойствам с самостоятельными глобулярными белками, называются модулями или доменами. Например, в дегидрогеназах два домена, один связывает НАД+ и этот домен сходен по строению у всех НАД-зависимых дегидрогеназ, а другой домен связывает субстрат и отличается по структуре у разных дегидрогеназ. Синтаза жирных кислот, представляющая одну полипептидную цепь, имеет 7 доменов, для катализа 7 реакций. Предполагается, что домены синтазы некогда объединились в один белок в результате слияния генов. Соединение модулей (доменов) в один белок способствует быстрому появлению и эволюции новых функциональных белков. КЛАССИФИКАЦИЯ БЕЛКОВ В организме человека содержится свыше 50 000 индивидуальных белков, отличающихся первичной структурой, конформацией, строением активного центра и функциями. Однако до настоящего времени нет единой и стройной классификации, учитывающей различные особенности белков. В основе имеющихся классификаций лежат разные признаки. Так белки можно классифицировать: · по форме белковых молекул (глобулярные – округлые или фибриллярные – нитевидные) · по молекулярной массе (низкомолекулярные, высокомолекулярные) · по выполняемым функциям (транспортные, структурные, защитные, регуляторные и др.) · по локализации в клетке (ядерные, цитоплазматические, лизосомальные и др.) · по структурным признакам и химическому составу белки делятся на две группы: простые и сложные. Простые белки представлены только полипептидной цепью, состоящей из аминокислот. Сложные белки имеют в своем составе белковую часть и небелковый компонент (простетическую группу). Однако и эта классификация не является идеальной, поскольку в чистом виде простые белки встречаются в организме редко.
Альбумины и глобулины. Альбумины (А) и глобулины (Г). А и Г белки, которые есть во всех тканях. Сыворотка крови наиболее богата этими белками. Содержание альбуминов в ней составляет 40-45 г/л, глобулинов 20-30 г/л, т.е на долю альбуминов приходится более половины белков плазмы крови. Альбумины -белки относительно небольшой молекулярной массы (15-70 тыс. Да); они имеют отрицательный заряд и кислые свойства, ИЭТ - 4,7, содержат много глутаминовой аминокислоты. Это сильно гидратированые белки, поэтому они осаждаются только при большой концентрации водоотнимающих веществ. Благодаря высокой гидрофильности, небольшим размерам молекул, значительной концентрации альбумины играют важную роль в поддержании осмотического давления крови. Если концентрация альбуминов ниже 30 г/л, изменяется осмотическое давление крови, что приводит к возникновению отеков. Около 75-80 % осмотического давления крови приходится на долю альбуминов. Характерным свойством альбуминов является их высокая адсорбционная способность. Они адсорбируют полярные и неполярные молекулы, выполняя транспортную роль. Это неспецифические переносчики они транспортируют гормоны, холестерол, билирубин, лекарственные вещества, ионы кальция. Связывание и перенос длинноцепочных жирных кислот - основная физиологическая функция сывороточных альбуминов. Альбумины синтезируются преимущественно в печени и быстро обновляются, период их полураспада 7 дней. Глобулины - белки с большей, чем альбумины молекулярной массой. Глобулины слабокислые или нейтральные белки (ИЭТ = 6 – 7,3). Некоторые из глобулинов обладают способностью к специфическому связыванию веществ (специфические переносчики). Возможно фракционирование белков сыворотки крови на альбумины и глобулины методом высаливания с помощью (NH4)2SO4. В насыщенном растворе осаждаются альбумины как более легкая фракция, в полунасыщенном – глобулины. В клинике широкое распространение получил метод фракционирования белков сыворотки крови путем электрофореза. При электрофоретическом разделении белков сыворотки крови можно выделить 5–7 фракций: Характер и степень изменения белковых фракций сыворотки крови при различных патологических состояниях представляет большой интерес для диагностических целей. Уменьшение альбуминов наблюдается в результате нарушения их синтеза, при дефиците пластического материала, нарушении синтетической функции печени, поражении почек. Содержание глобулинов увеличивается при хронических инфекционных процессах.
Электрофорез белков сыворотки крови.
Проламины и глютелины. Это группа растительных белков, которые содержатся исключительно в клейковине семян злаковых растений, где выполняют роль запасных белков. Характерной особенностью проламинов является то, что они не растворимы в воде, солевых растворах, щелочах, но растворимы в 70% растворе этанола, в то время как все другие белки выпадают в осадок. Наиболее изучены белки глиадин (пшеница) и зеин (кукуруза). Установлено, что проламины содержат 20-25% глутаминовой кислоты и 10-15 % пролина. Эти белки, например, глиадин, в норме у человека расщепляются, но иногда при рождении фермент, расщепляющий этот белок, отсутствует. Тогда этот белок превращается в продукты распада, обладающие токсическим действием. Развивается заболевание целиакия - непереносимость растительных белков. Глютелины – тоже растительные белки, не растворимые в воде, в растворах солей, этаноле. Они растворимы в слабых щелочах.
Протеиноиды. Белки опорных тканей (костей, хрящей, сухожилий, связок), кератины - белки волос, рогов, копыт, коллагены - белки соединительной ткани, эластин - белок эластических волокон. Все эти белки относятся к фибриллярным, не гидролизуются в желудочно - кишечном тракте. Коллаген составляет 25-33 % от общего количества белка организма взрослого человека или 6 % от массы тела. Пептидная цепь коллагена содержит около 1000 аминокислотных остатков, из которых каждая 3-я аминокислота – глицин, 20% составляют пролин и гидроксипролин, 10% аланин. При формировании вторичной и третичной структур этот белок не может давать типичных a-спиралей, поскольку аминокислоты пролин и оксипролин могут давать только одну водородную связь. Поэтому полипептидная цепь на участке, где находятся эти аминокислоты, легко изгибается, так как не удерживается, как обычно, второй водородной связью. Эластин – это основной структурный компонент эластических волокон, которые содержатся в тканях обладающих значительной эластичностью (кровеносные сосуды, связки, легкие). Свойства эластичности проявляются высокой растяжимостью этих тканей и быстрым восстановлением исходной формы и размера после снятия нагрузки. В составе эластина содержится много гидрофобных аминокислот (глицина, валина, аланина, лейцина, пролина).
СЛОЖНЫЕ БЕЛКИ Сложные белки кроме полипептидных цепей содержат в своем составе небелковую (простетическую) часть, представленную различными веществами. В зависимости от химической природы небелковой части выделяют следующие группы сложных белков:
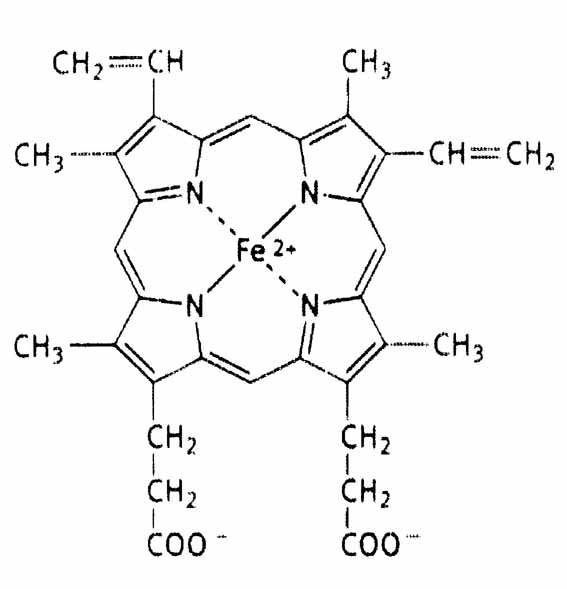
· хромопротеины · углевод – белковые комплексы · липид – белковые комплексы · нуклеопротеины · фосфопротеины ХРОМОПРОТЕИНЫ Хромопротеины – это сложные белки, простетическая часть которых представлена окрашенным компонентом (от греч. Chromos – краска). К ним относятся биологически важные белки гемоглобин, миоглобин, а также некоторые ферменты: каталаза, пероксидаза, цитохромы, все они являются гемпротеинами, так как простетическая часть их содержит гем. Гемоглобин (Нв). Гемоглобин имеет четвертичную структуру, молекулярная масса его 66-68 тыс.Да. Как следует из названия Нв представляет собой соединение гема с белком глобином. Это олигомерный белок, состоящий из 4 субъединиц (протомеров), соединенных нековалентными связями (гидрофобные, ионные взаимодействия, водородные связи). Субъединицы Нв «узнают» друг друга благодаря присутствию на их поверхности комплементарных по форме участков. Каждая из субъединиц или полипептидных цепей обозначается буквами, их 4 у Нв взрослого человека (Нв А), эти цепи называются альфа (a) и бета (b). Каждая молекула Нв А содержит по две a- и по две b-цепи. Они различаются первичной структурой и длиной полипептидной цепи: a - цепи содержат по 141 аминокислотному остатку, b - цепи по 146. Каждый из протомеров гемоглобина представляет собой природный координационный комплекс. В нем комплексообразователем служит Fe 2+. Его координационное число равно 6. Поэтому ион железа связан с 6 лигандами. 4 лиганда являются пиррольными кольцами протопорфирина. Пятая связь затрачивается на соединение с белком глобином через аминокислоту гистидин, а 6 координационная связь используется для связывания с кислородом или другими газообразными веществами (СО2, СО).
Строение гема. Внутри каждого протомера имеется гидрофобный "карман", в котором располагается гем, способный присоединять кислород.
Схема молекулы гемоглобина.
Основная функция гемоглобина – перенос кислорода из легких к периферическим тканям. Первая молекула кислорода изменяет конформацию протомера, к которому она присоединилась. Поскольку этот протомер соединен многими связями с другими протомерами, изменяется конформация и сродство других протомеров к лигандам. Это явление называют кооперативностью изменения конформации протомеров. Изменение конформации таковы, что сродство гемоглобина ко 2-й молекуле кислорода увеличивается. В свою очередь присоединение 2-ой, а затем и 3-ей молекулы кислорода, так же изменяет конформацию и облегчает присоединение следующих молекул кислорода. Сродство гемоглобина к 4-ой молекуле кислорода примерно в 300 раз больше чем к 1-ой.
Модель молекулы гемоглобина.
Кроме кислорода, молекула гемоглобина может быть связана с другими лигандами. Например, при соединении Hb с оксидом углерода (II) (угарный газ СО), образуется карбоксигемоглобин (НвСО). Причем гемоглобин имеет большее сродство к угарному газу чем к кислороду. Поэтому если в воздухе содержится угарный газ, гемоглобин легче с ним связывается и теряет способность связывать кислород. Наступает смерть от удушья, от недостаточного снабжения тканей кислородом. Возможно образование еще одного производного гемоглобина - карбгемоглобина, когда гемоглобин связывается с СО2. Однако СО2 связывается не с гемом, а присоединяется к NH2 – группам глобина. Образование карбгемоглобина используется для выведения СО2 из тканей к легким. Этим путем выводится 10-15% СО2. Типы гемоглобинов. Гемоглобины могут различаться по белковой части. Различают физиологические и аномальные типы гемоглобинов. Физиологические типы образуются на разных этапах нормального развития организма, а аномальные - вследствие нарушения последовательности аминокислот в белке глобине физиологических типов гемоглобина. Физиологические типы гемоглобинов отличаются друг от друга набором полипептидных цепей или субъединиц, образующихся на разных этапах развития организма человека – от эмбрионального до взрослого состояния. Различаются следующие физиологические типы гемоглобинов: а) примитивный НвР, появляется на самых ранних стадиях развития эмбриона (1 – 2 недели) Эмбриональный гемоглобин – тетрамер б) фетальный гемоглобин НвF (от лат. Fetus – плод). НвF является главным типом гемоглобина плода и составляет к моменту рождения 70% всего гемоглобина – это тетрамер ( в) гемоглобин взрослого НвА, НвА2, НвА3 (от лат. Adultus – взрослый). НвА появляется на более поздних стадиях развития плода, в крови взрослого человека примерно 95 – 96% НвА – это тетрамер (2?, 2?). Гемогловин А2 – тетрамер. Его содержание в эритроцитах взрослого человека равно 2%. Аномальные гемоглобины. Их обнаружено более 200 и отличаются они составом цепей или заменой аминокислот в полипептидных цепях. Из аномальных гемоглобинов часто встречается HвS или серповидно-клеточный Нв. Он обнаруживается у больных серповидно-клеточной анемией. Это широко распространенное заболевание в странах Южной Америки, Африки и Юго-Восточной Азии. При этой патологии эритроциты в условиях низкого парциального давления кислорода принимают форму серпа. Гемоглобин S отличается по ряду свойств от нормального гемоглобина. После отдачи кислорода в тканях он превращается в плохо растворимую форму и начинает выпадать в осадок в виде веретенообразных кристаллоидов. Последние деформируют клетку и приводят к массивному гемолизу. Химический дефект при серповидно-клеточной анемии сводится к замене в белке одной аминокислоты на другую. В норме в b - субъединицах тетрамерной структуры гемоглобина в шестом положении со стороны N – конца находится глутаминовая аминокислота, боковая группа которой имеет отрицательный заряд и характеризуется высокой гидрофильностью. При серповидно-клеточной анемии глутаминовая аминокислота заменена на гидрофобную аминокислоту – валин. Однако одной этой замены оказалось достаточно не только для нарушения формы эритроцита, но и для развития болезни. Миоглобин также относится к хромопротеинам. Это белок, имеющий третичную структуру. Вторичная и третичная структура миоглобина и протомеров гемоглобина очень сходны. Функции миоглобина и гемоглобина одинаковы. Оба белка участвуют в транспорте кислорода. Гемоглобин присоединяет кислород из альвеолярного воздуха и доставляет его в ткани. Миоглобин присоединяет кислород, доставленный гемоглобином и служит промежуточным звеном в транспорте кислорода внутри клетки к митохондриям, а также для запасания кислорода в тканях, создавая кислородный резерв, который расходуется по мере необходимости. В условиях интенсивной мышечной работы, когда парциальное давление кислорода в тканях падает, кислород освобождается из комплекса с миоглобином и используется в митохондриях клеток для получения энергии, необходимой для работы мышц.
ГЛИКОПРОТЕИНЫ (ГП) ПРОТЕОГЛИКАНЫ ГП - это сложные белки, содержащие олигосахаридные (гликановые) цепи, ковалентно связанные с белковой основой. К этой группе химических соединений относятся многие белки внешней поверхности цитоплазматических мембран и большинство секретируемых белков. ГП могут содержать до 40% углеводов, но как правило, в молекуле преобладает белковая часть. Короткие углеводные цепи ГП построены из глюкозамина, галактозамина, глюкозы, галактозы. Наиболее значимые моносахариды в составе ГП – N – ацетилглюкозамин, N – ацетилгалактозамин, N – ацетилнейраминовая (сиаловая) кислота. Число коротких углеводных цепей в ГП может доходить до 300-800. Длина и степень разветвленности углеводных цепей значительно варьирует. В составе одной олигосахаридной цепи находится не более 15-20 моносахаридных остатков. ГП присутствуют во всех классах белков – ферментах, гормонах, транспортных, структурных белках. Представители ГП – коллаген, эластин, иммуноглобулины.
Углеводный компонент, даже небольшой по массе, сообщает качественно новые свойства молекуле белка гликопротеинов. Для гликопротеинов характерна термостабильность, в отличие от простых белков ГП выдерживают высокие и низкие температуры без изменения физико-химических свойств. ГП в отличие от других белков с трудом перевариваются протеолитическими ферментами. Углеводная часть придает белку большую специфичность. Это своего рода векторные группы сложных белков, «узнающие» участки других структур (макромолекул, поверхности клеток). ГП быстрее выводятся из клетки и находятся, как правило, вне клетки. Гликопротеины выполняют следующие функции: Транспортная роль. ГП осуществляют транспорт гидрофобных веществ и ионов металлов. Так функцию переносчика железа выполняет ГП – трансферрин; меди – церуллоплазмин; стероидных гормонов – транскортин. Каталитическая. Углеводный компонент обнаружен в составе некоторых ферментов: энтерокиназа, пероксидаза, глюкозооксидаза, холинэстераза. Функция защитной смазки. Гликопротеины являются составными веществами муцинов слюны, желудочного и кишечного муцинов. ЛИПИД – БЕЛКОВЫЕ КОМПЛЕКСЫ Это комплексы белков с липидными компонентами, их условно подразделяют на две группы: Свободные липопротеины. Липопротеины плазмы крови, молока, растворимы в воде. Структурные протеолипиды. Входят в состав биомембран, растворимы в жирах. Липид – белковые комплексы в качестве небелковой части содержат липидные компоненты. Высшие жирные кислоты
Свободные липопротеины. Содержатся в плазме крови, все они имеют разную плотность (от 0,92 до 1,21 кг/л) благодаря липидному компоненту. В крови человека присутствуют несколько фракций ЛП, отличающихся по плотности, что связазано с различным соотношением липидного и белкового компонента в молекуле. ФРАКЦИИ ЛП: А) Хиломикроны (ХМ). Это самая низкая по плотности фракция, т.к. в составе их преобладают липидные комплексы и на долю белка приходится до 2 %. Плотность 0,95 кг/л. Хиломикроны появляются в сыворотке крови после приема жирной пищи; Б) Липопротеины очень низкой плотности (ЛПОНП). или пре-b-липопротеины, их плотность 0,94 – 1,006 кг/л; В) Липопротеины низкой плотности (ЛПНП) или b-липопротеины. Плотность их 1,006 – 1,063 кг/л; Г) Липопротеины высокой плотности (ЛПВП) или a-липопротеины. Плотность их 1,063 – 1,210 кг/л. В составе их преобладает белковый компонент. РОЛЬ СВОБОДНЫХ (ПЛАЗМЕННЫХ) ЛИПОПРОТЕИНОВ. Свободные ЛП играют транспортную роль, поэтому их называют - транспортными формами липидов. Благодаря своей растворимости в водной среде они могут переносить липиды, поступающие в кровь при всасывании из кишечника, а также распределять липиды между тканями, одни из которых их синтезируют, а другие используют. ЛП переносят триацилглицеролы, фосфолипиды, стероиды, а также небольшое количество жирорастворимых витаминов, b-каротина. В настоящее время доказана роль фракций липопротеинов в патогенезе атеросклероза – они называются атерогенными ЛП. К ним относятся ЛПНП и ЛПОНП, а фракцию ЛПВП называют антиатерогенной, так как её увеличение препятствует развитию атеросклероза. НУКЛЕОПРОТЕИНЫ Нуклепротеины – это сложные белки, небелковая часть которых представлена нуклеиновыми кислотами. Поскольку нуклеиновые кислоты бывают двух типов, нуклеопротеины делятся по составу на 2 группы: рибонуклеопротеины и дезоксирибонуклеопротеины. Нуклеиновые кислоты – это высокомолекулярные соединения, состоящие из мононуклеотидов, т.е. их структурной единицей является мононуклеотид (нуклеотид). Каждый нуклеотид включает 3 химически различных компонента: моносахарид, азотистое основание, остаток фосфорной кислоты. Нуклеотиды, входящие в РНК и ДНК, отличаются друг от друга по составу.
Соединение основания и пентозы называют нуклеозидом, связь между пентозой и азотистым основанием (b - гликозидная) образована - первым атомом углерода пентозы с первым атомом азота в пиримидиновых нуклеозидах и девятым атомом азота в пуриновых нуклеозидах. Нуклеотиды представляют собой соединения нуклеозидов с фосфорной кислотой (связь сложно – эфирная). В составе РНК и ДНК по 4 нуклеозидмонофосфата (нуклеотида). Вторичная структура ДНК. Особенностью нуклеотидного состава ДНК является то, что число адениловых нуклеотидов равно числу цитидиловых: А=Т, Г=Ц, следовательно, А+Г=Т+Ц, т.е. число пуриновых нуклеотидов равно числу пиримидиновых (правила Чаргаффа). Такие соотношения не свойственны РНК. Исходя из правил Чаргоффа о нуклеотидном составе ДНК и из рентгеноструктурных исследований, Дж. Уотсон и Ф. Крик (Великобритания) предложили модель строения ДНК (1953) Согласно этой модели молекула ДНК представляет собой двойную спираль, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотидные цепи в ней антипараллельны, т.е. если одна из них ориентирована в направлении 3' a 5', то вторая – в направлении 5' a 3'. Поэтому на каждом из концов молекулы ДНК расположены 5' – конец одной цепи и 3' – конец другой цепи. Все основания цепей ДНК (гидрофобные по свойствам) расположены внутри двойной спирали, а пентозы и остатки фосфорной кислоты – снаружи. Полинуклеотидные цепи удерживаются относительно друг друга за счет водородных связей, образующихся за счет специфического взаимодействия между парами комплементарных оснований. Комплементарными являются А и Т, они образуют две водородные связи, а также Г и Ц образуют три водородные связи. Кроме водородных связей в стабилизации вторичной структуры ДНК участвуют гидрофобные взаимодействия возникающие за счет гидрофобных азотистых оснований, обращенных внутрь спирали. Гидрофобные взаимодействия вносят основной вклад в стабилизацию двойной спирали, больший чем водородные связи между цепями. Рибозофосфатные связи располагаются по периферии, образуя ковалентный остов спирали
Особенности структуры РНК. Первичная структура РНК аналогична первичной структуре ДНК и представляет полинуклеотидную цепь, состоящую из мононуклеотидов, соединенных 3' a 5' – фосфодиэфирными связями. Вторичная структура РНК. Молекулы РНК построены из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные петли "шпильки", за счет водородных связей между комплементарными азотистыми основаниями А-У и Г-Ц. Участки цепи РНК в таких спирализованных участках антипараллельны, но не всегда полностью комплементарны. Наличие спирализованных участков характерно для всех типов РНК. Такие спирализованные участки содержат небольшое число нуклеотидных пар (до 20-30) и чередуются с неспирализованными участками. Основные типы РНК. По особенностям структуры и функциям различают 3 типа рибонуклеиновых кислот – транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомальные РНК (рРНК). Они различаются по первичной структуре, молекулярной массе, конформации, продолжительности жизни и по функциональной активности. 1. Транспортные РНК (тРНК) Пространственную структуру тРНК, независимо от различий в последовательности нуклеотидов, описывают универсальной моделью "клеверного листа". В каждой молекуле тРНК есть участки цепи, не участвующие в образовании водородных связей между нуклеотидными остатками. К ним в частности, относят участок, ответственный за связывание с аминокислотой на 3'-конце молекулы и антикодон – специфический триплет нуклеотидов, взаимодействующий комплементарно с кодоном мРНК. Транспортные РНК составляют около 15% всей РНК клетки, они участвуют в транспорте аминокислот.
Строение транспортных РНК.
1. Рибосомные РНК (рРНК) – компоненты рибосом. На долю рРНК приходится около 80% всей РНК клетки. 2. Матричные РНК (мРНК) составляют около 2% от всей РНК клетки. Матричные РНК называются также информационными РНК (иРНК). ФОСФОПРОТЕИНЫ. Фосфопротеины – это сложные белки, содержащие в своем составе в качестве простетической части фосфорную кислоту. Фосфорная кислота связана сложно – эфирной связью с белковой частью молекулы через гидроксильные группы оксиаминокислот (серин, треонин). Установлено, что фосфопротеины в клетках синтезируются в результате фосфорилирования при участии протеинкиназ. К фосфопротеинам относится казеиноген молока, который представляет собой белок с сильно выраженными гидрофильными свойствами. Казеиноген в молоке находится в виде кальциевой соли. Поэтому организм получает с молоком необходимые аминокислоты, кальций, лабильно связанный фосфор, находящиеся в казеиногене.
Производные моносахаридов Модификация имеющихся групп или введение новых заместителей в молекулу моносахаридов дает различные их производные. Они используются для построения разнообразных полимерных углеводов. Некоторые из производных являются промежуточными продуктами обмена.
Сиаловые кислоты во многом определяют взаимодействие лиганда с рецепторами клеточных мембран. Изменение содержания сиаловых кислот на клеточной поверхности сопровождает дифференцировку клеток и развитие патологического процесса. Избыточным количеством сиаловых кислот на поверхности мембран объясняют многие свойства опухолевых клеток, десиалирование рецепторов мембран гепатоцита приводит к развитию атеросклероза. Олигосахариды К олигосахаридам относятся сложные углеводы, имеющие от 2 до 10 звеньев моносахаридов соединенных гликозидными связями. Среди наиболее распространенных олигосахаридов следует отметить дисахариды – мальтозу, лактозу, сахарозу. Они отличются друг от друга составом моносахаридов, типом гликозидной связи, свойствами. Лактоза и мальтоза относятся к восстанавливающим дисахаридам. Гликозидная связь в них образуется за счет полуацетальной (гликозидной) ОН-группы одного моносахарида и спиртовой группой другого моносахарида. В этих дисахаридах имеется свободная полуацетальная окси-группа. Они обладают восстанавливающими свойствами, т.е. дают положительную реакцию Троммера с гидроксидом меди (II), что сопровождается образованием осадка кирпично-красного цвета (оксида меди-I-(Сu2О)). В отличие от мальтозы и лактозы сахароза относится к невосстанавливающим дисахаридам, т.к. гликозидная связь в молекуле сахарозы образуется за счет полуацетальных ОН-групп обоих моносахаридов. Поэтому сахароза не содержит свободного гликозидного гидроксила, она не может переходить в открытую карбонильную форму и поэтому не дает реакции на альдегидную группу (реакцию Троммера). Мальтоза (солодовый сахар) образуется при расщеплении крахмала в кишечнике. Содержится в больших количествах в солоде и солодовых экстрактах, отсюда и получила свое название "солодовый сахар". В растворе мальтоза может быть в открытой и циклической форме. Мальтоза состоит из остатков?, D-глюкопиранозы и D-глюкозы, связь между ними? (1> 4) гликозидная. Лактоза – молочный сахар; важнейший дисахарид молока. В коровьем молоке содержится до 5% лактозы, в женском молоке – до 8%. Лактоза состоит из?, D-галактопиранозы и D-глюкозы, связь?-1,4-гликозидная, поскольку в лактозе содержится свободный полуацетальный гидроксил, она относится к восстанавливающим сахарам Лактоза применяется в фармацевтической промышленности при изготовлении порошков и таблеток, т.к. она менее гигроскопична чем сахар
Сахароза - растворимый дисахарид сладкого вкуса. Содержится в сахарной свекле, сахарном тростнике. Сахароза не содержит свободного полуацетального гидроксила, поэтому относится к невосстанавливающим сахарам.
Гомополисахариды (ПС). Структурные различия между полисахаридами определяются: - строением моносахаридов, составляющих цепь - типом гликозидных связей, соединяющих мономеры в цепь - последовательностью остатков моносахаридов в цепи. В зависимости от строения остатков моносахаридов полисахариды делятся на гомополисахариды, макромолекулы которых состоят из моносахаридных остатков одного вида, гетерополисахариды содержат разные моносахариды. В зависимости от выполняемых функций ПС делят на 3 группы: - резервные ПС, выполняющие энергетическую роль. Эти ПС служат источником глюкозы. ПС менее растворимы чем моносахариды, следовательно они не влияют на осмотическое давление и поэтому могут накапливаться в клетке, например, крахмал – в клетках растений, гликоген – в клетках животных; - структурные ПС; - ПС, входящие в состав межклеточного матрикса, принимают участие в образовании тканей, а также в пролиферации и дифференцировке клеток. ПС межклеточного матрикса водорастворимы и сильно гидратированы. К гомополисахаридам относятся крахмал, гликоген, клетчатка.
|
||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 257; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.221.53.209 (0.132 с.) |