Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Вопрос 1. Анатомо-физиологические особенности системы крови.Содержание книги
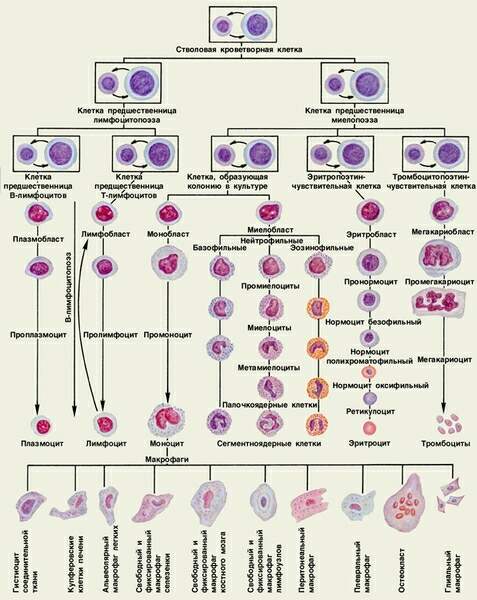
Поиск на нашем сайте Существуют несколько теорий кроветворения, но в настоящее время общепринятой является унитарная теория кроветворения, на основании которой была разработана схема кроветворения (И. Л. Чертков и А. И. Воробьев, 1973 г.).
В процессе поэтапной дифференцировки стволовых клеток в зрелые форменные элементы крови в каждом ряду кроветворения образуются промежуточные типы клеток, которые в схеме кроветворения составляют классы клеток. Всего в схеме кроветворения различают 6 классов клеток: 1класс-стволовые клетки; Морфологическая и функциональная характеристика клеток различных классов схемы кроветворения. 1 класс - стволовая полипотентная клетка, способная к поддержанию своей популяции. По морфологии соответствует малому лимфоциту, является полипотентной, то есть способной дифференцироваться в любой форменный элемент крови. Направление дифференцировки стволовой клетки определяется уровнем содержания в крови данного форменного элемента, а также влиянием микроокружения стволовых клеток - индуктивным влиянием стромальных клеток костного мозга или другого кроветворного органа. Поддержание численности популяции стволовых клеток обеспечивается тем, что после митоза стволовой клетки одна из дочерних клеток становится на путь дифференцировки, а другая принимает морфологию малого лимфоцита и является стволовой. Делятся стволовые клетки редко (1 раз в полгода), 80 % стволовых клеток находятся в состоянии покоя и только 20 % в митозе и последующей дифференцировке. В процессе пролиферации каждая стволовая клетка образует группу или клон клеток и потому стволовые клетки в литературе нередко называются клон-образующие единицы - КОЕ. 2 класс - полустволовые, ограниченно полипотентные (или частично коммитированные) клетки - предшественницы миелопоэза и лимфопоэза. Имеют морфологию малого лимфоцита. Каждая из них дает клон клеток, но только миелоидных или лимфоидных. Делятся они чаще (через 3-4 недели) и также поддерживают численность своей популяции. 3 класс - унипотентные поэтин-чувствительные клетки - предшественницы своего ряда кроветворения. Морфология их также соответствует малому лимфоциту. Способны дифференцироваться только в один тип форменного элемента. Делятся часто, но потомки этих клеток одни вступают на путь дифференцировки, а другие сохраняют численность популяции данного класса. Частота деления этих клеток и способность дифференцироваться дальше зависит от содержания в крови особых биологически активных веществ - поэтинов, специфичных для каждого ряда кроветворения (эритропоэтины, тромбопоэтины и другие). Первые три класса клеток объединяются в класс морфологически неидентифицируемых клеток, так как все они имеют морфологию малого лимфоцита, но потенции их к развитию различны. 4 класс - бластные (молодые) клетки или бласты (эритробласты, лимфобласты и так далее). Отличаются по морфологии как от трех предшествующих, так и последующих классов клеток. Эти клетки крупные, имеют крупное рыхлое (эухроматин) ядро с 2-4 ядрышками, цитоплазма базофильна за счет большого числа свободных рибосом. Часто делятся, но дочерние клетки все вступают на путь дальнейшей дифференцировки. По цитохимическим свойствам можно идентифицировать бласты разных рядов кроветворения. 5 класс - класс созревающих клеток, характерных для своего ряда кроветворения. В этом классе может быть несколько разновидностей переходных клеток - от одной (пролимфоцит, промоноцит), до пяти в эритроцитарном ряду. Некоторые созревающие клетки в небольшом количестве могут попадать в периферическую кровь (например, ретикулоциты, юные и палочкоядерные гранулоциты). 6 класс - зрелые форменные элементы крови. Однако следует отметить, что только эритроциты, тромбоциты и сегментоядерные гранулоциты являются зрелыми конечными дифференцированными клетками или их фрагментами. Моноциты не окончательно дифференцированные клетки. Покидая кровеносное русло, они дифференцируются в конечные клетки - макрофаги. Лимфоциты при встрече с антигенами, превращаются в бласты и снова делятся. Совокупность клеток, составляющих линию дифференцировки стволовой клетки в определенный форменный элемент, образуют его дифферон или гистологический ряд. Например, эритроцитарный дифферон составляет:
В процессе созревания эритроцитов в 5 классе происходит: синтез и накопление гемоглобина, редукция органелл, редукция ядра. В норме пополнение эритроцитов осуществляется в основном за счет деления и дифференцировки созревающих клеток пронормоцитов, базофильных и полихроматофильных нормоцитов. Такой тип кроветворения носит название гомопластического кроветворения. При выраженной кровопотери пополнение эритроцитов обеспечивается не только усиленным делением созревающих клеток, но и клеток 4, 3, 2 и даже 1 классов гетеропластический тип кроветворения, предшествующий собой уже репаративную регенерацию крови.Кровь представляет собой жидкость (жидкая ткань мезодермального происхождения), красного цвета, слабо щелочной реакции, солоноватого вкуса с удельным весом 1,054-1,066. Совместно с тканевой жидкостью и лимфой она образует внутреннюю среду организма. Кровь выполняет многообразные функции. Главнейшие из них следующие: - транспорт питательных веществ от пищеварительного тракта к тканям, местам резервных запасов от них (трофическая функция); - транспорт конечных продуктов метаболизма из тканей к органам выделения (экскреторная функция); - транспорт газов (кислорода и диоксида углерода из дыхательных органов к тканям и обратно; запасание кислорода (дыхательная функция); - транспорт гормонов от желез внутренней секреции к органам (гуморальная регуляция); - защитная функция - осуществляется за счет фагоцитарной активности лейкоцитов (клеточный иммунитет), выработки лимфоцитами антител, обезвреживающих генетически чужеродные вещества (гуморальный иммунитет); - свертывание крови, препятствующее кровопотере; - терморегуляторная функция - перераспределение тепла между органами, регуляция теплоотдачи через кожу; - механическая функция - придание тургорного напряжения органам за счет прилива к ним крови; обеспечение ультрафильтрации в капиллярах капсул нефрона почек и др.; - гомеостатическая функция - поддержание постоянства внутренней среды организма, пригодной для клеток в отношении ионного состава, концентрации водородных ионов и др. Относительное постоянство состава и свойств крови - гомеостаз является необходимым и обязательным условием жизнедеятельности всех тканей организма. Из всего объёма крови примерно половина циркулирует по организму. Остальная же половина задерживается в расширенных капиллярах некоторых органов и называется депонированной. Органы, в которых депонирована кровь, называются кровяным депо. Схема кроветворения (И. Л. Чертков и А. И. Воробьев, 1973 г.).
Селезёнка. Вмещает в своих лакунах - отростках капилляров до 16% всей крови. Эта кровь практически выключена из кругооборота и не смешивается с циркулирующей кровью. При сокращении гладких мышц селезёнки лакуны сжимаются, и кровь поступает в общее русло. Печень. Вмещает в себя до 20% объёма крови. Печень выполняет роль кровяного депо за счёт сокращения сфинктеров печёночных вен, по которым кровь оттекает от печени. Тогда в печень крови поступает больше, чем оттекает. Капилляры печени расширяются, кровоток в ней замедляется. Однако депонированная в печени кровь полностью не выключается из кровотока. Подкожная клетчатка. Депонирует до 10% крови. В кровеносных капиллярах кожи имеются анастомозы. Часть капилляров расширяется, заполняется кровью, а кровоток совершается по укороченным путям (шунтам). Лёгкие также можно отнести к органам, депонирующим кровь. Объём сосудистого русла лёгких также не постоянен, он зависит от вентиляции альвеол, величины кровяного давления в них и от кровенаполнения сосудов большого круга кровообращения. Таким образом, депонированная кровь выключена из кровотока и в основном не смешивается с циркулирующей кровью. Вследствие всасывания воды депонированная кровь более густа, она содержит большее количество форменных элементов.Значение депонированной крови заключается в следующем. Когда организм находится в состоянии физиологического покоя, его органы и ткани не нуждаются в усиленном снабжении кровью. В этом случае депонирование крови снижает нагрузку на сердце, и в результате оно работает на 1/5 - 1/6 своей мощности. При необходимости кровь может быстро перейти в кровоток, например при физической работе, сильных эмоциональных переживаниях, вдыхании воздуха с повышенным содержанием диоксида углерода - то есть во всех случаях, когда требуется, увеличит доставку кислорода и питательных веществ органам. В механизмах перераспределения крови между депонированной и циркулирующей участвует вегетативная нервная система: симпатические нервы вызывают увеличение объёма циркулирующей крови, а парасимпатические - переход крови в депо. При поступлении в кровь большого количества адреналина происходит выход крови из депо. При кровопотерях объём крови восстанавливается, прежде всего, за счёт перехода тканевой жидкости в кровь, а затем в кровоток поступает депонированная кровь. В результате объём плазмы восстанавливается значительно быстрее, чем количество форменных элементов. При увеличении объёма крови (например, при введении большого количества кровезаменителей или при выпаивании большого количества воды) часть жидкости быстро выводится почками, но большая часть переходит в ткани, а затем постепенно выводится из организма. Таким образом, восстанавливается объём крови, заполняющий сосудистое русло. Метаболизм и обмен железа У взрослого здорового человека в среднем содержится около 3-4 г железа (40-50 мг Fe/кг массы тела). Около 60 % (2,4 г) всего железа находится в гемоглобине, а примерно 30 % железа входит в состав ферритина - депо железа. Депо железа - величина непостоянная, определяемая разницей между поступившим и выделенным из организма железом. Около 9 % железа находится в миоглобине, белке, переносящем кислород в мышцах. Приблизительно 1 % железа входит в состав ферментов, таких как цитохромы, каталазы, пероксидазы и др. Железо всасывается как в виде гема (10 % поглощаемого железа), так и в негемовой (90 %) форме с помощью ворсинок верхней части тонкого кишечника. Сбалансированная ежедневная диета содержит около 5-10 мг железа (гемового и негемового), но всасывается лишь 1-2 мг. Гемовое железо содержится лишь в небольшой части пищевого рациона (мясные продукты). Оно очень хорошо всасывается (на 20-30 %) и на его усвоение не влияют другие компоненты пищи. Большая часть пищевого железа – негемовое. В клетках слизистой оболочки тонкого кишечника, во время процесса всасывания, закисное железо (Fe(II)) превращается в окисное железо (Fe(III)) для того, чтобы быть включенным в состав трансферрина и транспортироваться по всему организму. Трансферрин синтезируется печенью. Он отвечает за транспортировку не только всосавшегося в кишечнике железа, но и железа, поступающего из разрушенных эритроцитов для повторного использования. В физиологических условиях заняты не более, чем 30 % железосвязывающих рецепторов трансферрина плазмы. Несвязанное с белками железо токсично, так как ион Fe+++ запускает реакции свободно-радикального окисления, повреждающие клеточные структуры. Основной источник железа для кроветворения - это распадающийся в РЭС гемоглобин старых эритроцитов (гем идет на реутилизацию, а глобин распадается), а железо пищи является лишь дополнительным источником.Железо хранится в организме в виде ферритина и гемосидерина. Из этих двух белков, на долю ферритина приходится большая часть хранимого железа, которое представлено в виде гидроокиси/окиси железа, заключенной в белковую оболочку, апоферритин. Существует 5 изоформ ферритина: щелочные изоформы печени и селезенки отвечают за депонирование железа, а кислые изоформы миокарда, плаценты, опухолевых клеток являются посредниками в процессах синтеза и участвуют в регуляции Т-клеточного иммунного ответа. Поэтому ферритин является еще и индикатором острого воспаления и опухолевого роста. Снижение ферритина ниже 15 мкг/л является достоверным показателем истинного дефицита железа. Наиболее богаты ферритином предшественники эритроцитов в костном мозге, макрофаги и ретикулоэндотелиальные клетки печени. Гемосидерин рассматривают как уменьшенную форму ферритина, в которой молекулы потеряли часть их белковой оболочки и сгруппировались вместе. При избытке железа, часть его, хранимая в печени в виде гемосидерина, увеличивается. Запасы железа расходуются и возмещаются медленно и, поэтому, недоступны для экстренного синтеза гемоглобина при компенсации последствий острого кровотечения или других видов кровопотерь. Гемосидерин – это нерастворимое производное ферритина, форма депонирования избыточного железа, откладывающегося в тканях в виде зерен. Гемосидерин медленно мобилизуется из тканей, может приводить к повреждению клеток паренхиматозных органов (гемосидерозу). Выведение железа в норме в количестве 1 мг/сутки происходит с калом, слущенным эпителием кожи и слизистых; у женщин во время месячных в количестве до 15 мг в сутки. При распаде старых эритроцитов в селезенке железо гема не теряется, а в виде комплекса с трансферрином направляется в органы кроветворения на реутилизацию. В случае внутрисосудистого гемолиза свободный гемоглобин сберегается от потерь через почки путем связывания с гаптоглобином плазмы в крупномолекулярный комплекс. При массивном гемолизе запасы гаптоглобина быстро истощаются, и свободный гемоглобин теряется с мочой. Витамин В12 – это комплексное металлоорганическое соединение, в котором атом кобальта соединен с циклом коррина. Витамин В12 не синтезируется в организме и должен поступать с пищей.
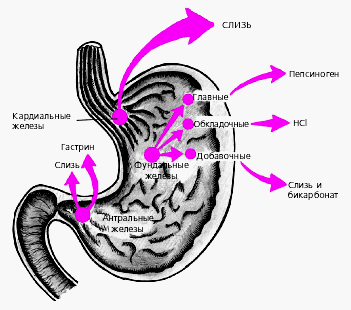
В желудке витамин В12, содержащейся в пище, высвобождается и образует стойкое соединение желудочным с R-протеином, и поступает в двенадцатиперстную кишку, где и расщепляется; В12 освободившись от протеина, соединяется с внутренним фактором Кастла. Внутренний фактор Кастла представляет собой гликопротеид с молекулярной массой 50 000. Его секретируют обкладочные клетки желудка вместе с соляной кислотой. Комплекс витамин В12 с внутренним фактором Кастла доходит до дистального отдела подвздошной кишки, где связывается с рецепторами на щеточной каёмке и поглащается энтероцитами. Здесь внутренний фактор Кастла разрушается, а витамин В12 связывается с белком транскоболамином II, поступает в кровь и поглащается печенью, костным мозгом и другими тканями
|
||||||
|
|
Последнее изменение этой страницы: 2016-12-13; просмотров: 392; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.113 (0.013 с.) |