Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Пресинаптическое нервное окончание.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
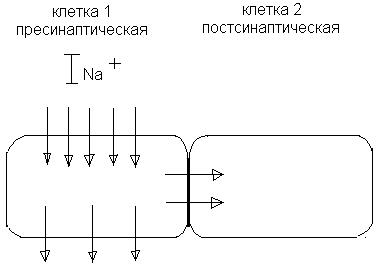
Характерно наличие большого количества субмикроскопических структур округлой формы, которые называют синаптическими пузырьками (везикулами), имеются митохондрии. Синаптическая щель. Ширина 10 – 50 нм (100-500 А). При таких размерах электрическая передача возбуждения практически невозможна из-за значительной потери тока во внеклеточной среде, поэтому химическая передача возбуждения представляет собой необходимый усиливающий механизм. Постсинаптическая мембрана (ПСМ). Малое количество электроуправляемых натриевых каналов, а потому низкая чувствительность к электрическому току. Следовательно, невозможность генерировать ПД. Зато имеются специфические хемочувствительные рецептороуправляемые каналы. Под влиянием медиатора изменяется проницаемость для ионов K+ и Na+, в результате чего изменяется мембранный потенциал. В зависимости от природы медиатора и свойств рецепторов ПСМ может происходить деполяризация мембраны, что характерно для возбуждения, или гиперполяризация, что типично для торможения. В случае деполяризации ПСМ говорят о возникновении возбуждающего постсинаптического потенциала (ВПСП), а при гиперполяризации – о тормозящем постсинаптическом потенциале (ТПСП). В состоянии покоя некоторые везикулы с медиатором подходят к пресинаптической мембране (ПреСМ) и медиатор попадает в синаптическую щель, диффундирует, вступает во взаимодействие с рецепторами ПСМ и обусловливает возникновение миниатюрных потенциалов. При возбуждении пресинаптического окончания взаимосвязь между деполяризацией ПреСМ и высвобождением медиатора обеспечивают ионы Са+, которые поступают снаружи в нервное окончание. Механизм секреции медиатора регулируется рядом биологически активных веществ, в том числе самими медиаторами, а также циклическими нуклеотидами и нейропептидами, которые являются в данном случае модуляторами синаптической передачи. Основные этапы синаптической передачи. Приход ПД к пресинаптической мембране, ее деполяризация и генерация на ней потенциала действия. Проникновение внутрь пресинаптической мембраны ионов кальция – для транспорта везикул с медиатором. Взаимодействие везикул (синаптических пузырьков) с активными участками пресинаптической мембраны. Экзоцитоз и выделение квантов медиатора в синаптическую щель (квант медиатора – это содержимое одной везикулы; квант ацетилхолина примерно 400 – 40000 молекул). Диффузия медиатора к постсинаптической мембране. Взаимодействие медиатора с клеточными рецепторами ПСМ. Изменение проницаемости для ионов. Образование постсинаптических потенциалов. Синапсы и рецепторы постсинаптических мембран этих синапсов могут подразделяться в зависимости от того, какой медиатор выделяется в синаптическую щель. Например: 1.Холинергические (ацетилхолин – АЦХ) 5 – 10% всех синапсов. Н–холинергические рецепторы (никотинзависимые) в одних случаях (нервно-мышечные синапсы) блокируются курареподобными веществами, в других (синапсы головного мозга, синапсы вегетативной нервной системы) не реагируют на курареподобные вещества, но блокируются ганглиоблокаторами (бензогексоний и ему подобные вещества). М–холинергические рецепторы (мускаринзависимые) имеются в центральных холинергических синапсах, а также в окончаниях постганглионарных парасимпатических волокон. Все М-ХР блокируются атропином. В отличие от Н-ХР синапсов, в М-ХР синапсах может иметь место не только возбуждение, но и торможение. Часть М-ХР синапсов являются возбуждающими (гладкомышечные клетки (ГМК) желудочно-кишечного тракта и бронхов), а часть – тормозными (в сердечной мышце). 2. Адренергические. 0,5% всех синапсов. Медиатор – норадреналин (НА). Эффект выделения НА зависит от того, какой вид рецептора находится на ПСМ. Различают 4 вида адренорецепторов (АР). Их делят на класс альфа-АР (внутри класса 2 популяции: альфа-1 и альфа-2) и бета-АР (соответственно бета-1 и бета-2). В каждом синапсе есть, вероятно, все 4 вида рецепторов, но доминирует всегда какой-то один. При взаимодействии НА с альфа-1-АР происходит деполяризация ПСМ (образуется ВПСМ). Альфа-1-АР много в ГМК сосудов кожи и желудочно-кишечного тракта, в нейронах головного мозга. Альфа-2-АР находятся на ПреСМ адренергического синапса, благодаря чему НА оказывает тормозной антидромный эффект (тормозит выделение очередной порции НА). Бета-1-АР в основном представлены в миокарде, благодаря чему НА вызывает здесь активацию. Бета-2-АР в основном расположены в ГМК сосудов скелетных мышц, коронаров, в ГМК бронхов, матки. При их активации возникает торможение активности соответствующих структур. Т.о. возбуждающий или тормозный характер действия медиатора определяется не столько медиатором, сколько свойствами постсинаптической мембраны, находящимися на ней рецепторами. В ЦНС есть синапсы, медиаторами которых могут быть: серотонин (0,5% всех синапсов); дофамин; гистамин; АТФ; глицин; ГАМК (25-40% синапсов). Глицин, ГАМК – в тормозных синапсах ЦНС. Инактивация медиатора. Необходима для реполяризации ПСМ, восстановления исходного потенциала. Например, АЦХ действует на рецепторы ПСМ нервно-мышечного синапса 1-2 мс. Потом часть АЦХ диффундирует в лимфу и кровь, а часть гидролизуется ферментом – ацетилхолинэстеразой на холин и уксусную кислоту, которые снова поступают в пресинаптическое окончание и являются продуктами для синтеза новых порций медиатора. Для норадреналина ингибитором является моноаминооксидаза и катехолоксиметилтрансфераза. Но для норадреналина наиболее характерен «обратный захват» пресинаптическими структурами. Несмотря на наличие ингибиторов НА разрушается ими в незначительном количестве и снова депонируется синаптическими пузырьками. Свойства химических синапсов. 1. Возбуждение проводится в одном направлении. 2. Количество выделяющегося медиатора пропорционально частоте приходящей нервной импульсации. В покое: 1 квант в 1 сек; при ПД – 200 квантов за 2-3 мс. 3. Синаптическая передача не подчиняется закону «все или ничего». Возможна суммация ПСП на ПСМ. 4. Трансформация ритма. Скорость проведения возбуждения в синапсе меньше, чем по нерву. Синаптическая задержка (около 0,5 мс). 5. Высокая чувствительность к химическим веществам, недостатку кислорода. 6. Высокая утомляемость. Развивается в результате длительного высокочастотного стимулирования. Обусловлено истощением и несвоевременным синтезом медиатора в пресинаптической части синапса или глубокой, стойкой деполяризацией ПСМ (пессимальное торможение). Электрическая передача. Электрические синапсы. Встречаются редко. ПД вызывает возбуждение в соседней клетке или торможение без химического посредника. Концептуально были предсказаны еще до открытия химических синапсов (назывались эфапсами). Между 1930 – 1950 г.г., когда концепция химической синаптической передачи была уже общепризнанной, выяснилось, что межклеточная передача возбуждения может осуществляться и электрическим способом. Принцип: 2 соседние клетки прилегают тесно друг к другу (рис. 12). Сопротивление электрическому току соприкасающихся мембран должно быть сравнимо с сопротивлением остальной, внесинаптической области мембраны. При возбуждении клетки 1 натриевый ток входит в нее через открытые натриевые каналы и выходит через пока не возбужденные участки мембраны. Часть тока входит через участок мембранного контакта в клетке 2, вызывая ее деполяризацию. Уровень деполяризации здесь примерно в 10 раз ниже, чем в клетке 1, однако выше порога генерирования ПД в клетке 2. Часто такая деполяризация подпороговая, а клетка 2 возбуждается в результате суммации синаптических потенциалов.
Рис. 12. Электрический синапс.
Ионы, переносящие электрические токи не могут проходить через липидные мембраны. Для их транспорта в «мембранных контактах» между электрически сопряженными клетками необходимы канальные белки. Такие межклеточные связи называются нексусами, или «щелевыми контактами». Щелевые контакты соединяют группы синхронно функционирующих клеток. В тканях не относящихся к НС, клетки также бывают соединены щелевыми контактами. В миокарде и гладкой мускулатуре эти контакты создают функциональный синцитий. Возбуждение здесь переходит от одной клетки к другой без заметной паузы или снижения амплитуды ПД на границе. Для таких органов важна регулируемость щелевых контактов. Их каналы закрываются при снижении рН или повышении концентрации Са2+. Это происходит в случае повреждения клеток или глубокого нарушения обмена. За счет такого механизма пораженные места изолируются от остальной части функционального синцития, и распространение патологии ограничивается (например, при инфаркте миокарда). Кроме этих возбудимых тканей существует и много других, где клетки также соединены щелевыми контактами (все эпителии, печень). В принципе такая связь присуща любой клетке на ранних стадиях эмбрионального развития, когда все клетки соединены между собой щелевыми контактами и сохраняют их до стадии дифференцировки органов. Щелевой контакт – наиболее распространеный тип электрического синапса. Однако существуют и другие. Например, электрическим путем может передаваться и торможение. В этом случае ПД особым образом расположенных пресинаптических волокон генерируют во внеклеточном пространстве вокруг постсинаптического аксона местный положительный потенциал такой амплитуды, что его деполяризация не может достичь порогового уровня, и проведение по нему ПД блокируется. Учитывая широкое распространение щелевых контактов, кажется удивительным, почему в нервной системе они не используются для синаптической передачи повсеместно. Видимо, сложнее организованные химические синапсы обеспечивают настолько более высокую специфичность и регулируемость межклеточной коммуникации, что в значительной степени вытеснили электрические.
Эфаптическая передача. При некоторых заболеваниях аксоны теряют свою миелиновую оболочку, становясь демиелинизированными. Демиелинизированные аксоны могут вступать в аномальные взаимодействия, когда импульсы, проходящие по группам нервных волокон, индуцируют возбуждение других параллельно идущих аксонов. Это называется эфаптической передачей. Когда такие аномальные ПД генерируются в сенсорных нервных волокнах, появляются аномальные ощущения, парестезии. Если они связаны с ноцицептивными (болевыми) волокнами, то возникают такие неприятные синдромы, как невралгия, каузалгия, невромные боли. Межаксонные помехи могут быть следствием не только недостаточной изоляции (миелиновыми оболочками), но и повышенной возбудимости аксонов.
|
||||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 660; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.148.108.174 (0.01 с.) |