
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Классификация нервных центров.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
По локализации в структурах нервной системы различают корковые, подкорковые и спинальные центры. В головном мозге также выделяют центры диэнцефальные, мезэнцефальные, бульбарные, гипоталамические, таламические. На функциональной основе центры нервной системы разделяют по регулируемой функции (например, сосудодвигательный центр, центр теплообразования, дыхательный центр и др.) или по афферентному восприятию (например, центры зрения, слуха, обоняния и др.). Выделяют также центры нервной системы, которые формируют мотивационные состояния организма, являясь пейсмекерами мотивационных возбуждений (центры голода, жажды, насыщения и др.). Существуют центры нервной системы, которые на основе интеграции возбуждений формируют целостные реакции организма (например, центры глотания, чиханья, дефекации, половой центр и т.п.). Тонус нервных центров. От центров нервной системы идет больший или меньший поток импульсов к эффекторам, обусловливая их постоянное тоническое возбуждение. Тоническое состояние центра нервной системы определяется соотношением активированных и так называемых молчащих нервных клеток, что обеспечивается не только поступающими от периферических рецепторов афферентными импульсами, но и гуморальными влияниями (гормоны, метаболиты, биологически активные вещества). Доминанта. При высоком уровне тонического возбуждения центр может доминировать в деятельности ЦНС (принцип доминанты А.А. Ухтомского). В доминантном центре нервной системы нейроны характеризуются длительной повышенной возбудимостью, возрастает эффективность временной и пространственной суммации возбуждений. Доминирующий центр, подавляя деятельность других центров, может стать главенствующим в формировании целенаправленного поведенческого акта животных и человека. Примером доминанты служит классический опыт А.А. Ухтомского, в котором раздражение сенсомоторной коры на фоне механического раздувания прямой кишки вызывало реакцию дефекации вместо сокращения лапы животного. Линейное распространение возбуждения. Наиболее жесткая линейная связь осуществляется афферентными проекционными путями, обеспечивающими передачу возбуждений от периферических специализированных рецепторов через различные ядра головного мозга вплоть до коры большого мозга. Возбуждение от рецепторов поступает в задние столбы спинного мозга, затем через ядра продолговатого мозга и вентробазальный комплекс таламуса достигает соматосенсорной области коры большого мозга. Генерализованное распространение возбуждения. -обеспечивает функциональное взаимодействие между корой большого мозга и подкорковыми структурами. Эффект восходящих генерализованных активирующих влияний ретикулярных образований проявляется в возбуждении всех областей коры большого мозга, что находит отражение в десинхронизации суммарной биоэлектрической активности мозга. В головном мозге существуют нисходящие, главным образом кортико-фугальные влияния на подкорковые структуры. Эти структуры могут вновь активировать кору мозга восходящими потоками импульсов, создавая эффект возвратной генерализации возбуждений. Взаимодействие восходящих и нисходящих влияний обусловливает двустороннюю связь между структурами головного мозга, особенно между корой мозга и подкорковыми образованиями — реверберацию возбуждений.
#10 Дайте современное объяснение структуры связей нейронов и мембранных процессов, лежащих в основе известных видов центрального торможения. Объясните результаты опытов И.М.Сеченова и Гольца с современных позиций. Механизмы торможения проявляются в прекращении или уменьшении активности нервных клеток. В отличие от возбуждения торможение — локальный нераспространяющийся процесс, возникающий на клеточной мембране. Сеченовское торможение. Наличие процесса торможения в ЦНС впервые было показано Сеченовым в 1862 г. в экспериментах на лягушке. Выполняли разрез головного мозга лягушки на уровне зрительных бугров и измеряли время рефлекса отдергивания задней лапы при погружении ее в раствор серной кислоты (метод Тюрка). При наложении на разрез зрительных бугров кристаллика поваренной соли время рефлекса увеличивалось. Прекращение воздействия соли на зрительные бугры приводило к восстановлению исходного времени рефлекторной реакции. Рефлекс отдергивания лапки обусловлен возбуждением спинальных центров. Кристаллик соли, раздражая зрительные бугры, вызывает возбуждение, которое распространяется к спинальным центрам и тормозит их деятельность. И.М. Сеченов пришел к выводу, что торможение является следствием взаимодействия двух и более возбуждений на нейронах ЦНС. В этом случае одно возбуждение неизбежно становится тормозимым, а другое — тормозящим. Подавление одним возбуждением другого происходит как на уровне постсинаптических мембран (постсинаптическое торможение), так и за счет уменьшения эффективности действия возбуждающих синапсов на пресинаптическом уровне (пресинаптическое торможение). Пресинаптическое торможение. Пресинаптическое торможение развивается в пресинаптической части синапса за счет воздействия на его мембрану аксо-аксональных синапсов. В результате как деполяризующего, так и гиперполяризующего воздействия происходит блокирование проведения импульсов возбуждения по пресинаптическим путям к постсинаптической нервной клетке. Постсинаптическое торможение. Наибольшее распространение в ЦНС имеет механизм постсинаптического торможения, которое осуществляется специальными тормозными вставочными нервными клетками (например, клетки Реншоу в спинном мозге или клетки Пуркинье (грушевидные нейроны) в коре мозжечка). Особенность тормозных нервных клеток состоит в том, что в их синапсах имеются медиаторы, вызывающие на постсинаптической мембране нейрона ТПСП (тормозящие постсинаптические потенциалы), т.е. кратковременную гиперполяризацию. Например, для мотонейронов спинного мозга гиперполяризующим медиатором является аминокислота глицин, а для многих нейронов коры большого мозга таким медиатором служит гамма-аминомасляная кислота — ГАМК. Частным случаем постсинаптического является возвратное торможение. Реципрокное торможение. Механизм постсинаптического торможения лежит в основе таких видов торможения, как реципрокное и латеральное. Реципрокное торможение является одним из физиологических механизмов координации деятельности нервных центров. Так, попеременно реципрокно тормозятся в продолговатом мозге центры вдоха и выдоха, прессорный и депрессорный сосудодвигательные центры. Реципрокное торможение проявляется на уровне спинного мозга при осуществлении строго координированных двигательных актов (ходьба, бег, чесание). На уровне сегментов спинного мозга возбуждение группы мотонейронов, вызывающих сокращение мышц-сгибателей, сопровождается реципрокным торможением другой группы мотонейронов, приводящих к расслаблению мышц-разгибателей. Латеральное торможение. Активность нейронов или рецепторов, расположенных рядом с возбужденными нейронами или рецепторами, прекращается. Механизм латерального торможения обеспечивает дискриминаторную способность анализаторов. Так, в слуховом анализаторе латеральное торможение обеспечивает различение частоты звуков, в зрительном анализаторе латеральное торможение резко увеличивает контрастность контуров воспринимаемого изображения, а в тактильном анализаторе способствует дифференцировке двух точек прикосновения. При поступлении возбуждений к синапсам нервной клетки на постси-наптических мембранах могут возникать процессы гиперполяризации. Гиперполяризация приводит к возрастанию критического уровня деполяризации мембраны, следовательно, затрудняет возникновение возбуждения. Такие постсинаптические потенциалы получили название «тормозящие постсинаптические потенциалы» (ТПСП); они возникают в синапсах, где медиатор вызывает гиперполяризацию постсинаптической мембраны. Каждый нейрон синтезирует в своем теле и затем выделяет во всех своих синапсах один и тот же медиатор, поэтому нейроны и ацетилхолиновой передачей возбуждения называются холинергическими, с адреналиновой – адренергическими. К гиперполяризующим медиаторам относят ГАМК глицин. Эти медиаторы взаимодействуя с хеморецепторами постсинаптической мембраны, приводят к развитию ТПСП.
#11 Охарактеризуйте основные методы исследования ЦНС (электроэнцефалография, импульсная активность нейронов), объясните их использование для оценки функционального состояния чел-ка. Методы исследования функций ЦНС делятся на две группы: 1) непосредственное изучение и 2) опосредованное (косвенное) изучение. Методы непосредственного изучения функций ЦНС подразделяют на морфологические и функциональные. Морфологические методы. К морфологическим методам относятся мак-роанатомическое и микроскопическое исследования строения мозга. Этот принцип лежит в основе метода генетического картирования мозга, позволяющего выявлять функции генов в метаболизме нейронов. К морфологическим методам относят и метод меченых атомов. Сущность его заключается в том, что вводимые в организм радиоактивные вещества интенсивнее проникают в те нервные клетки мозга, которые в данный момент наиболее функционально активны. Функциональные методы: разрушение и раздражение структур ЦНС, стереотаксический метод, электрофизиологические методы. Метод разрушения. Разрушение структур мозга является довольно грубым методом исследования, поскольку повреждаются обширные участки мозговой ткани. Так, повреждение в области продолговатого мозга приводит к нарушению дыхания, глотания, деятельности сердца и изменению тонуса сосудов. В клинике для диагностики повреждений мозга различного происхождения (опухоли, инсульт и др.) у человека используют методы компьютерной рентгенотомографии, эхоэнцефалографии, ядерного магнитного резонанса. Метод раздражения структур мозга позволяет установить пути распространения возбуждения от места раздражения к органу или ткани, функция которых при этом изменяется. В качестве раздражающего фактора чаще всего применяют электрический ток. В эксперименте на животных применяют метод самораздражения различных участков мозга: животное получает возможность посылать раздражение в мозг, замыкая цепь электрического тока и прекращать раздражение, размыкая цепь. Стереотаксический метод введения электродов. Стереотаксические атласы, которые имеют три координатных значения для всех структур мозга, помещенного в пространство трех взаимно перпендикулярных плоскостей — горизонтальной, сагиттальной и фронтальной. Данный метод позволяет не только с высокой точностью вводить электроды в мозг с экспериментальной и диагностической целями, но и направленно воздействовать на отдельные структуры ультразвуком, лазерными или рентгеновскими лучами с лечебной целью, а также проводить нейрохирургические операции. Электрофизиологические методы исследования ЦНС включают анализ как пассивных, так и активных электрических свойств мозга. Электроэнцефалография. Метод регистрации суммарной электрической активности мозга называется электроэнцефалографией, а кривая изменений биопотенциалов мозга — электроэнцефалограммой (ЭЭГ). ЭЭГ регистрируют с помощью электродов, располагаемых на поверхности головы человека. Используют два способа регистрации биопотенциалов: биполярный и монополярный. При биполярном способе регистрируют разность электрических потенциалов между двумя близко расположенными точками на поверхности головы. При монополярном способе регистрируют разность электрических потенциалов между любой точкой на поверхности головы и индифферентной точкой на голове, собственный потенциал которой близок к нулю. Такими точками являются мочки уха, кончик носа, а также поверхность щек. Основными показателями, характеризующими ЭЭГ, являются частота и амплитуда колебаний биопотенциалов, а также фаза и форма колебаний. По частоте и амплитуде колебаний различают несколько видов ритмов в ЭЭГ. Гамма >35 Гц, эмоциональное возбуждение, умственная и физическая деятельность, при нанесении раздражения. Бета 13-30 Гц, эмоциональное возбуждение, умственная и физическая деятельность, при нанесении раздражения. Альфа 8-13 Гц состояние умственного и физического покоя, с закрытыми глазами. Тета 4-8 Гц, сон, умеренные гипоксии, наркоз. Дельта 0,5 – 3,5 глубокий сон, наркоз, гипоксия. Основным и наиболее характерным ритмом является альфа-ритм. В состоянии относительного покоя альфа-ритм наиболее выражен в затылочных, затылочно-височных и затылочно-теменных областях головного мозга. При кратковременном действии раздражителей, например света или звука, появляется бета-ритм. Бета- и гамма-ритмы отражают активированное состояние структур головного мозга, тета-ритм чаще связан с эмоциональным состоянием организма. Дельта-ритм указывает на снижение функционального уровня коры большого мозга, связанное, например, с состоянием легкого сна или утомлением. Локальное появление дельта-ритма в какой-либо области коры мозга указывает на наличие в ней патологического очага. Микроэлектродный метод. Регистрация электрических процессов в отдельных нервных клетках. Микроэлектроды - стеклянные или металлические. Стеклянные микропипетки заполняют раствором электролита, чаще всего концентрированным раствором хлорида натрия или калия. Существуют два способа регистрации клеточной электрической активности: внутриклеточный и внеклеточный. При внутриклеточном расположении микроэлектрода регистрируют мембранный потенциал, или потенциал покоя нейрона, постсинаптические потенциалы — возбуждающий и тормозящий, а также потенциал действия. Внеклеточный микроэлектрод регистрирует только положительную часть потенциала действия.
#12 Объясните характер взаимодействия нейронов сегментов спинного мозга и проприорецепторов опорно-двигательного аппарата в мех-х поддержания мышечного тонуса. Интегративная деятельность спинного мозга связана со структурной организацией его сегментов. Каждый сегмент спинного мозга получает афферентную импульсацию через задние (чувствительные) корешки и посылает возбуждение к эффекторам через передние (двигательные) корешки. Таким образом, уже в пределах каждого сегмента спинного мозга возможна интеграция простейших двигательных реакций. Миотатический рефлекс — одна из простых реакций на растяжение мышцы, осуществляемая на уровне спинного мозга по самому короткому пути — от проприорецепторов до мотонейронов и далее к двигательным мышечным волокнам. Реакция растяжения является основой регуляции длины мышцы, изменение которой возникает при поднятии груза или при сокращении мышцы-антагониста. Скелетные мышцы всегда находятся в состоянии некоторого напряжения. Постоянное незначительное напряжение мышц, не сопровождающееся признаками утомления, называется мышечным тонусом. Односторонняя перерезка у спинальной лягушки, подвешенной на крючке штатива, чувствительных (задних) корешков спинного мозга, в которых проходят афферентные нервные волокна, иннервирующие соответствующую заднюю лапку, приводит к исчезновению мышечного тонуса этой лапки и она распрямляется. К аналогичному эффекту приводит перерезка передних (двигательных) корешков или разрушение спинного мозга. Эти опыты свидетельствуют о том, что при разрушении основных звеньев рефлекторного кольца (афферентных и эфферентных путей, нервных центров) мышечный тонус исчезает. Следовательно, мышечный тонус имеет рефлекторную природу. Источником возбуждений, поддерживающих мышечный тонус, являются проприорецепторы. В скелетных мышцах имеются три вида проприорецепторов: • мышечные веретена, расположенные среди мышечных волокон; • сухожильные рецепторы Гольджи, расположенные в сухожилиях; • пачиниевы тельца, расположенные в фасциях, сухожилиях, связках. Особое значение в регуляции мышечного тонуса имеют мышечные веретена и сухожильные рецепторы Гольджи. Мышечные веретена представляют собой небольшие продолговатые образования, напоминающие своим внешним видом прядильные капсулы мышечного веретена находится пучок мышечных волокон, которые называются интрафузальными, т. к. они расположенными внутри веретена в отличие от обычных мышечных волокон, которые называются зкстрафузальными. Каждое интрафузальное волокно состоит из трех частей: • его центральная часть называется ядерной сумкой, в которой находятся ядра мышечной клетки; • два периферических участка, которые имеют поперечную исчерченность и обладают способностью сокращаться; • миотрубки, расположенные между ядерной сумкой и периферическими участками. Ядерную сумку в виде спирали окружают нервные волокна чувствительного нейрона- первичные рецепторные окончания. В области миотрубок нервные окончания афферентных нейронов гроздевидно ветвятся, образуя вторичные рецепторные окончания. В мышце мышечное веретено одним концом прикрепляется к экстрафузальному мышечному волокну, а другим - к сухожилию этого волокна. Таким образом, мышечное веретено расположено в мышце параллельно экстрафузальным мышечным волокнам. При снижении тонуса экстрафузального волокна увеличивается его длина, что приводит к растяжению и раздражению первичных и вторичных рецепторных окончаний, для которых растяжение является адекватным раздражителем. Возбуждение от рецепторных окончаний по афферентным волокнам поступает в спинной мозг к мотонейронам, расположенным в передних рогах. Мотонейроны спинного мозга принято подразделять на альфа- и гамма-мотонейроны (так как их аксоны относятся к А-альфа и А-гамма нервным волокнам). Возбуждение от альфа-мотонейронов поступает к экстрафузальным мышечным волокнам, вызывая их сокращение - тонус восстанавливается. Избыточное сокращение экстрафузальных мышечных волокон приводит к растяжению сухожильных рецепторов Гольджи, так как они прикрепляются к мышце последовательно. В них возникает возбуждение, которое поступает к тормозным вставочным нейронам спинного мозга, а от них к альфа-мотонейронам. Активность альфа-мотонейронов при этом снижается, уменьшается импульсация, идущая от них к экстрафузальным мышечным волокнам, тонус несколько снижается. Рассмотренные выше механизмы поддержания мышечного тонуса осуществляются на уровне спинного мозга, поэтому такой тонус называется спинальным или простейшим. Спинальный тонус характеризуется очень слабой выраженностью тонического напряжения. Такой тонус не может обеспечить поддержание позы животного и акт ходьбы, но он достаточен для осуществления простейших спинальных рефлексов. Большое значение в регуляции мышечного тонуса имеют базальные ядра - бледный шар и полосатое тело, которые образуют стриопаллидарную систему. Эти структуры регулируют активность всех нижележащих отделов ЦНС, участвующих в регуляции мышечного тонуса, обеспечивая адекватное перераспределение тонуса мышц при различных видах деятельности. При поражении экстрапирамидной системы, составной частью которой являются базальные ядра, возникают нарушения регуляций тонуса мускулатуры, что приводит к развитию так называемых дрожательных параличей. Главную роль в приспособительной регуляции мышечного тонуса выполняет кора головного мозга. С ее участием и участием других структур, регулирующих мышечный тонус, формируется нормальный или корковый тонус. Регуляция тонуса скелетной мускулатуры осуществляется экстрапирамидной системой головного мозга. Быстрые (фазные) движения обеспечиваются активностью пирамидной системы.
#13 Характер взаимодействия различных отделов ЦНС в процессах формирования позы. В стволе мозга (продолговатый, мост, средний) заложены цельные движения (в отличие от спинальных элементов движений) - стереотипные, врожденные, позные. Позные рефлексы направлены на 1. перемену и сохранение позы, 2. восстановление естественной позы при ее нарушении, 3. удержание позы при внезапных воздействиях, например, резком ускорении. Классификация позных рефлексов: Статические рефлексы: 1. позно-тонические - перемена и сохранение позы (наклон головы вправо приводит к повышению тонуса разгибателей правых конечностей), 2. выпрямительные - восстановление позы (переворот на конечности из положения лежа на спине). Статокинетические - удержание позы и равновесия при внезапных воздействиях (лифтный рефлекс - увеличение тонуса разгибателей при движении вверх, перераспределение мышечного тонуса при резком ускорении, например, в вагоне метро). Основные рефлексогенные зоны, с которых вызываются эти рефлексы - вестибулярный аппарат и проприорецепторы шеи, поэтому особо выделяют лабиринтные и шейные рефлексы. Однако они могут вызываться и с проприорецепторов туловища и конечностей, кожных рецепторов и т.д. Через спинной мозг или черепные нервы ствол мозга получает всю чувствительность, кроме обонятельной. Непосредственно (через черепные нервы) или через синной мозг ствол мозга управляет всеми структурами организма. Ядра ретикулярной формации и ретикулоспинальный путь: сохранение вертикального положения; вестибулярное ядро Дейтериса и вестибулоспинальный путь (вест. аппарат, проприорецепторы мм. шеи) - система равновесия; красное ядро и руброспинальный путь - система стереотипных движений (описано), четверохолмие и тектоспинальный путь - система настораживания. Основные входы ствола мозга: от коры (префронтальная извилина - моторная зона) - запускает стволовые движения, от мозжечка - корректирует стволовые движения.
#14. Характер взаимодействия различных отделов ЦНС при выполнении произвольного движения. Отделы коры головного мозга: 1. моторная зона (прецентральная извилина) - управляет отдельными группами мышц, непосредственно или через стволовые ядра (каждому отделу моторной коры соотв. своя группа мышц), 2. премоторная зона и дополнительная моторная зона (хранят программы наиболее частых автоматизированных приобретенных движений - мимики, речи, письма, бытовых навыков..), 3. префронтальная кора (замысел и планирование цельных поведенческих актов). После планирования двигательного акта сигналы идут к стволовым ядрам (особенно красному ядру, если движения стереотипные врожденные), или к вторичным двигательным зонам (например, произнесение слова), а далее к соотв. зонам моторной зоны. Остальное см. в #13.
#15. Явление функциональной ассиметрии мозга, ее значение при формировании поведения. Первичные моторные, соматосенсорные и зрительные зоны обслуживают только одну противоположенную зону тела, тогда как сложные поведенческие акты требуют участия всего организма в целом. Т.е., хотя вторичные (первичная зона - всегда получение инфы, вторичная зона - обработка) и ассоциативные (объединение нескольких типов информации и зон мозга) зоны имеются в каждом полушарии, одна половина подчиняет другую в отдельный момент времени. У большинства ведущее полушарие - левое, т.к. деятельность в основном связана с обработкой вербальных функций, в т.ч. абстрактно-логического мышления. В соотв. зонах правого полушария развиваются функции, отсутствующие в левом: интерпретация мимики, жестов и интонаций, восприятие образов и музыки. Оба полушария постоянно обмениваются информацией, а ведущую роль попеременно берет то одно, то другое.
#16. На основе знания принципов работы ЦНС объясните зависимость характера ответной реакции организма от места действия раздражителя и его параметров. Проведите сравнительный анализ соматических и вегетативных рефлексов, участвующих в регуляции различных физиологических функций. Хз чо говорить точно, но надо поболтать про различные виды типа: воспринимаются зрительным, слуховым, тактильным, вестибулярным, обонятельным, вкусовым анализаторами. И чем сильнее раздражитель, тем сильнее будет ответная реакция. Длительность раздражения и градиент тоже будут влиять. Силараздражения – з ависимость ответной реакции от силы раздражителя. · Пороговые (воспринимаются организмом) · Подпороговые (не воспринимаются) Вследствие этого ответная реакция будет иметь место или нет. Величина ответа прямо пропорциональна физической силе раздражителя. Длительность. Если раздражитель слабый, то бы как длительно он ни действовал, ответная реакция не возникает. Градиент. Тут надо сказать что-то про то, что Еслипостепенно увеличивать силу раздражения, то возникает привыкание.
Про рефлексы. По отвечающему органу: различают: • соматические, или двигательные, (рефлексы скелетных мышц), например флексорные, экстензорные, локомоторные, статокинетические и др.; • вегетативные (внутренних органов) — пищеварительные, сердечно-сосудистые, выделительные, секреторные и др. #17 Охарактеризуйте структурно-функциональные особенности и важнейшие физиологические свойства симпатического отдела ВНС. Симпатический отдел. Тела первых нейронов симпатического отдела ВНС расположены преимущественно в задних ядрах гипоталамуса, среднем и продолговатом мозге и в передних рогах спинного мозга, начиная с 1-го грудного и кончая 3—4-м сегментом поясничного ее отдела. Периферическая (исполнительная) часть ВНС представлена преганглионарными и ганглионарными нейронами. Симпатический отдел. Тела преганглионарных нейронов располагаются в сером веществе боковых рогов спинного мозга, начиная от 1-го грудного сегмента до 2—3-го поясничного сегмента, и в двигательных ядрах черепных нервов. Миелиновые аксоны этих нейронов представлены медленнопроводящими возбуждение В-волокнами. Аксоны преганглионарных нейронов через синапсы взаимодействуют с нейронами, расположенными в периферических ганглиях. Околопозвоночные ганглии расположены по обе стороны вдоль позвоночника в виде цепочек, составляющих правый и левый симпатические стволы. Продольно ганглии соединяются межузловыми соединительными ветвями, состоящими из миелиновых и безмиелиновых нервных волокон. Имеются и поперечные связи между ганглиями правой и левой стороны. Кроме этого, ганглии симпатического ствола соединяются со спинномозговыми нервами, благодаря чему они получают возможность иннервировать скелетную мускулатуру. Другая часть постганглионарных волокон направляется к внутренним органам. Особенности строения симпатических и парасимпатических нервов. Симпатические нервы сформированы в основном преганглионарными, а парасимпатические — постганглионарными нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими безмиелиновыми волокнами типа С. Локализация ядер – заднее ядро гипоталамуса, нейроны боковых рогов грудных и поясничных сегментов спинного мозга. Характеристика отростков – прерываются в ганглиях, преганглионарные короткие, тип В, постганглионарные длинные, тип С. Зона иннервации – иннервирует все внутренние органы Локализация – парабертебральные (пограничный столб), превертебральные (чревное, солнечное сплетение, брыжеечные узлы). Медиатор – ацетилхолин, норадреналин. Физиологические свойства симпатической нервной системы: 1.благодаря мультипликации в симпатических ганглиях распространяющееся из них возбуждение широко охватывает сразу несколько различных органов, т.е. является генерализованным. Эти влияния наиболее отчетливо прослеживаются при эмоциональных реакциях. 2.оказывает активирующие влиянияна функции иннервируемых органов: усиливает катаболические реакции, силу и частоту сокращений сердца, повышает артериальное давление, улучшает оксигенацию тканей, увеличивает содержания глюкозы в крови, скорость проведения возбуждения в скелетных мышцах и их тонус, расширяет бронхи, увеличивает объем легочной вентиляции; расширяет зрачки, увеличивает секрецию катехоламинов надпочечниками. При этом одновременно снижается тонус пищеварительного тракта, ослабляются процессы всасывания и ферментативного расщепления в кишечнике. 3. участвует в формировании целостных состояний, как агрессия, стресс, болевые реакции. 4. передача с пре- на ганглионарные нейроны осуществляется с помощью ацетилхолина, а на эффекторы — норадреналина. 5. представлена на периферии на значительном протяжении постганглионарными волокнами типа С, возбуждение на пути от ганглиев к органам распространяется медленнее, чем в парасимпатической системе. 6. Эффекты действия более продолжительны. 7. Электрические потенциалы в симпатических ганглиях характеризуются продолжительными следовыми явлениями. При стимуляции преганглионарных волокон регистрируется быстрый возбуждающий постсинаптический потенциал, который сменяется медленным тормозным постсинаптическим потенциалом. 8. начальная деполяризация обусловлена действием ацетилхолина на никотиновые рецепторы.
#18 Охарактеризуйте структурно-функциональные особенности и важнейшие физиологические свойства парасимпатического отдела ВНС. Парасимпатический отдел Центральные нейроны расположены в передних отделах гипоталамуса, среднем и продолговатом мозге, во 2-4-м сегментах крестцового отдела спинного мозга. Периферическая (исполнительная) часть ВНС представлена преганглионарными и ганглионарными нейронами. Парасимпатический отдел. Тела преганглионарных нейронов расположены в среднем и продолговатом мозге среди двигательных нейронов черепных нервов: глазодвигательного, лицевого, языкоглоточного и блуждающего, а также в сером веществе боковых рогов крестцового отдела спинного мозга. Вместе с черепными нервами преганглионарные волокна распространяются к ганглионарным нейронам, расположенным в крылонебном, подчелюстном и ушном ганглиях. Отходящие от них постганглионарные волокна иннервируют глазное яблоко, аккомодационную мышцу и сфинктер зрачка, поднижнечелюстную, подъязычную и околоушную слюнные железы. Преганглионарные волокна блуждающего нерва распространяются к нейронам, расположенным в ганглиях органов грудной и верхней части брюшной полости. Аксоны нейронов крестцового отдела спинного мозга через тазовые нервы адресуются к нейронам, располагающимся в ганглиях органов нижней части брюшной полости и таза, иннервируя нижнюю часть толстой кишки, прямую кишку, мочевой пузырь, нижнюю часть мочеточника и наружные половые органы. Особенности строения симпатических и парасимпатических нервов. Симпатические нервы сформированы в основном преганглионарными, а парасимпатические — постганглионарными нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими безмиелиновыми волокнами типа С. Локализация ядер – передний гипоталамус, средний мозг, мост, продолговатый мозг, боковые рога I-V крестцовых сегментов спинного мозга. Характеристика отростков -прерываются в ганглиях, преганглионарные длинные, тип В, постганглионарные короткие, тип С. Зона иннервации – ограничена (нет в надпочечниках и стенках большинства сосудов) Локализация –в иннервируемых органах (интрамурально) или рядом с ними. Медиатор -ацетилхолин Физиологические свойства парасимпатической нервной системы: 1. влияния направлены и локальны. Оказывает ограниченное воздействие в пределах иннервируемого органа. 2. оказывает успокаивающее, расслабляющее действие на большинство функций организма; снижается возбудимость ЦНС и миокарда, уменьшаются интенсивность метаболизма, сила и частота сердечных сокращений, кровяное давление, объем легочной вентиляции, температура тела; увеличивается секреция инсулина и как следствие снижается концентрация глюкозы в крови и увеличивается внутриклеточное депонирование ее в виде гликогена. При этом одновременно усиливаются моторная, секреторная и всасывательная функции желудочно-кишечного тракта. 3. усиливает анаболические реакции. 4.Парасимпатические влияния доминируют в формировании сна и психологического субъективного чувства удовлетворения. 5.Медиатором в преганглионарных и постганглионарных волокнах служит ацетилхолин. 6.Парасимпатическая нервная система представлена на значительном расстоянии преганглионарными волокнами типа В. Вследствие этого возбуждения от центра до органа доходят быстрее, чем по симпатическим нервам. 7.Эффекты действия парасимпатических нервов менее продолжительны. #19. Вегетативные рефлексы, имеющие клиническое значение. Рефлекс глазосердечный (Даньини-Ашнера): замедление пульса на 4 —8 ударов в 1 мин. и снижение артериального давления при надавливании на глазные яблоки; большее замедление указывает на повышенную возбудимость парасимпатической части вегетативной нервной системы. Рефлекс солярный: замедление пульса и снижение артериального давления при надавливании на брюшную стенку между пупком и мечевидным отростком (область чревного сплетения). Рефлекс синокаротидный: общее название P., возникающих при раздражении рецепторов, расположенных в сонном синусе. Прессорный синокаротидный рефлекс сопровождается повышением периферического сопротивления, давления в аорте, желудочках сердца, но частота сокращений сердца может не изменяться и сопротивление коронарных сосудов несколько возрастает. Геринга рефлекс: замедление пульса при задержке дыхания на стадии глубокого вдоха; если в положении сидя это замедление превышает 6 ударов в 1 мин., то оно свидетельствует о повышенной возбудимости блуждающего нерва.
#20. Оценивать вегетативный статус человека (индекс Кердо). Вегетативная реактивность: холодовая проба, ортопроба. Индекс Кердо — показатель, использующийся для оценки деятельности вегетативной нервной системы. Вычисляется по формуле
Index — Индекс Кердо DAD — Диастолическое артериальное давление Pulse — Пульс Если значение этого индекса больше нуля, то говорят о преобладании возбуждающих влияний в деятельности вегетативной нервной системы, если меньше нуля, то о преобладании тормозных, если равен нулю, то это говорит о функциональном равновесии.
Холодовая проба. Сущность холодовой пробы заключается в том, что при опускании предплечья в холодную воду (+4°С) происходит рефлекторное сужение артериол и артериальное давление повышается, причем, тем больше, чем больше возбудимость сосудодвигательных центров. В состоянии покоя у испытуемого на плечевой артерии трижды до получения стабильных цифр измеряют АД. Затем ему предлагают на 1 мин погрузить кисть правой руки (немного выше лучезапястного сустава) в воду температуры +4°С. АД измеряют сразу после прекращения холодового воздействия, а затем в начале каждой минуты в течение первых 5 мин восстановления и через каждые 3 мин последующего периода до момента регистрации АД, соответствующего исхо
|
||||
|
|
Последнее изменение этой страницы: 2016-08-26; просмотров: 438; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.138.134.77 (0.015 с.) |

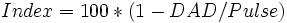 , где:
, где:


