Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Общая физиология возбудимых тканей.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
2-1. Произойдет деполяризация мембраны и снижение мембранного потенциала.
2-2. Мембранный потенциал увеличится (гиперполяриза-ция), так как калиевый ток теперь не будет уменьшаться за счет противоположного тока натрия, как было до опыта.
2-3. В этом случае выключается Na-K-насос, поляризация мембраны постепенно исчезнет, так как произойдет уравнивание концентраций натрия и калия по обе стороны мембраны
2-4. Если время действия раздражающего тока будет очень коротким (см. кривую Гоорвега-Вейса).
2-5. Возбуждение возникает при условии, если мембранный потенциал меньше или равен критическому уровню деполяризации. Поэтому в данном случае распрост-раняющееся возбуждение возникнет только в том случае, если мембранный потенциал уменьшится на величину, боль-шую чем 27 мв. (на 30%).
2-6. Возбудимость уменьшится, так как в этом случае необходимы большая сила и большее время, чтобы сдвинуть мембранный потенциал до критического уровня.
2-7. В данном случае новый мембранный потенциал стал равным 108 мв, а критический уровень деполяризации - 78 мв. Начальные величины этих показателей - 90 мв и 60 мв. Следовательно, исходная разница между мембранным потенциалом и критическим уровнем деполяризации не изменилась и осталась равной 30 мв. Это значит, что возбудимость данной мембраны не изменилась.
2-8. В данном случае исходная возбудимость мембраны соответствует разнице Ео и Ек в 30 мв. В начале деполяри-зации, когда мембранный потенциал был равен 90 мв, а разница Ео и Ек = 20 мв, возбуди-мость возросла на одну треть. После длительного раздражения критический уровень деполяризации достиг величины 54 мв. Так как в этом случае разница Ео и Ек составила 34 мв, то ясно, что возбу-димость ткани упала. Это явление носит название «катодической депресии» Вериго.
2-9. Мембранный потенциал под анодом увеличивается, а при выключении тока возвращается к исходному уровню. Следовательно, чтобы при размыкании под анодом могло возникнуть возбуждение, необходимо возрастание критического уровня деполяризации на такую величину, чтобы он стал равным исходному мемб-ранному потенциалу. Этот сдвиг не зависит от величины гиперполяризации, а определяется главным образом ее длительностью. Необходимый сдвиг равен 100-70=30 мв.
2-10. При замыкании мышца сократится, так как порог замыкательного удара посто-янного тока меньше размыкательного. При размыкании сокращения не будет.
2-11. Сокращения не будет, так как в этом случае катод находится ближе к мышце, и при сильном токе под ним возникает торможение (катодическая депрессия Вериго), которое блокирует возбуждение, приходящее от анода при размыкании тока.
2-12. Катод, так как возбуждение при замыкании возникает под катодом и ему ничто не мешает дойти до мышцы.
2-13. Катод. При размыкании катодическая депрессия блокирует движение воз-буждения от анода.
2-14. Схема возникновения возбуждения и последующего восстановления исходного состояния мембраны: Раздражение --- повышение возбудимости для Na --- вход Na внутрь клетки --- деполяризация --- генерация потенциала действия и перезарядка мембраны --- возникновение натриевой инактивации --- увеличение калиевой проницаемости --- фаза реполяризации --- активация K-Na- АТФ-азы --- включение K-Na-насоса --- восстановление мембранного потенциала.
2-15. В данном случае Ек = 70 мв. Так как возбуждение возникает при условии, если мембранный потенциал достиг кри-тического уровня деполяризации, то реобаза должна быть равной (100-70):5 = 6 мв.
2-16. Параметры возбудимости в этом случае будут следующими: реобаза - 0,8 вольт, хронаксия - 28 мсек.
2-17. Представлена цепь процессов, возникающих под катодом при действии пос-тоянного тока на ткань. Последователь-ность извращена. Правильно так: Пассивная деполяризация --- повышение натриевой проницаемости --- усиление потока натрия в клетку --- активная деполяризация --- локальный ответ --- потенциал действия.
2-18. Наименьший порог отмечается при прямоугольном стимуле раздражающего тока, так как при медленном нарастании тока из-за развития явлений аккомодации увеличивается пороговая сила.
2-19. При отведении потенциала первым способом регистрируется двуфазный ток действия, при втором способе - однофазный.
2-20. Уменьшение мембранного потенциала; деполяризация, местный потенциал, локальный ответ.
2-21. Потенциал покоя уменьшится, так как градиент концентрации калия будет меньше. Степень уменьшения - около 30%.
2-22. Наибольшая лабильность у нерва, наименьшая - у синапса.
2-23. А- нейрон (сома), Б - аксон, В - постсинаптическая мембрана, Г - мембрана мышечного волокна. Повышенной хими-ческой чувствительностью обладает постсинаптическая мембрана.
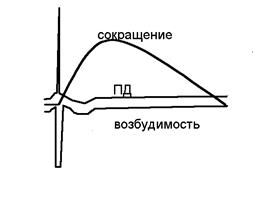
2-24. Схема соотношения потенциала действия, кривой возбудимости и кривой сокращения скелетной мышцы выглядит так:
2-25. Раздражение называется непрямым, если оно производится через нерв, подхо-дящий к мышце. От момента раздражения нерва до момента начала мышечного сок-ращения происходят следующие события: Возбуждение нерва --- движение возбуждения по нерву --- возбуждение преси-наптической мембраны ---- выделение медиатора --- возбуждение постсинапти-ческой мембраны --- возбуждение мемб-раны мышечного волокна --- движение возбуждения по мышечному волокну --- электро-механическое сопряжение --- активация акто-миозинового комплекса -- сокращение.
2-26. Неправильно, надо так:
2-27. Первая кривая - утомление нормальной мышцы. Вторая может быть получена при нарушении ресинтеза АТФ в клетке, при котором работа Са-насоса прекращается. Са++ накапливается в межфибриллярном пространстве и вызывает появление контрактуры. Мышца фиксируется в состо-янии укорочения.
2-28. Процесс передачи возбуждения в нервно-мышечном синапсе: Нервный импульс --- высвобождение ацетилхолина --- взаимодействие ацетил-холина с холинорецептором постсинаптической мембраны --- повышение ионной проницаемости ПСМ ---возникновение возбуждающего постсинаптического потенциала (ВПСП) --- возникновение ПД в мембране мышечного волокна и распро-странение его по волокну.
2-29. Для гладкого тетанического сокращения необходимо, чтобы интервал между раздражениями был длиннее реф-рактерного периода, но короче всей длительности сокращения. В данном случае этот интервал лежит в пределах от 10 до 70 мсек, значит при частоте от 15 до 100 гц будет наблюдаться тетанус. При мень-шей частоте будут одиночные сокращения, при большей - пессимум.
2-30. Так как скорость проведения волны сокращения совпадает со скоростью возбуждения, то в данном случае волна сокращения пройдет по всему волокну за 0,02 сек.
2-31. Так как длительность ПД совпадет с периодом рефрактерности, после которой следует фаза супернормальной возбуди-мости, интервал между раздражениями должен быть 10 мсек. Это соответствует частоте раздражения 100 гц, Чтобы импульс попал в субнормальный период, необходимо применение более частых раздражений.
2-32. Приведена схема электромеханического сопряжения: Раздражение --- возникновение ПД --- проведение его вдоль клеточной мембраны вглубь волокна по Т-системе --освобождение Са++ из саркоплазматического ретикулюма --- взаимодействие актина и миозина --- сокращение мышечного волокна – активация Са-насоса --- возвращение Са++ в цистерны ретикулюма --- расслабление мышцы.
2-33. Мышца снова начнет сокращаться, так как при раздражении нервно-мышечного препарата утомление раньше всего наступает в синапсе.
2-34. Мышца расслабится, так как атропин блокирует передачу импульсов в адрен-ергических синапсах.
2-35. Если весь Са++ возвращается в ретикулюм за 100 мсек, значит суммация сокращения и зубчатый тетанус будут возникать при частоте больше 10 гц. При частоте раздражения 50 гц перерыв между импульсами в 5 раз короче, и за это время в ретикулюм вернется уже не 1 функциональная единица Са++, а только 1/5 единицы. 4/5 же остаются в межфибриллярном пространстве и накапливаются там. Поскольку максимальная концентрация Са+ (5*10 6 мэкв/л) в 10 раз больше критической (0,5*106 мэкв/л), то такое количество Са++ накопится в пространстве через 10:4/5=12,5 импульсов. Это значит, что в ответ на 13 импульс мышца даст максимальную высоту сокращения.
2-36. В данном случае теоретически ткань могла бы воспроизводить1000: 5 =200 импульсов. В условии сказано, что истинная лабильность в 4 раза меньше, т.е. равна 50 гц. Значит, при частоте разд-ражения 10 гц мышца будет отвечать одиночными сокращениями или зубчатым тетанусом, при 50 гц - гладким, а при часто те более 50 гц возникнет пессимум частоты.
2-37. При частоте немного меньше 150 гц. Лабильность синапса - 149 гц, так как это максимальная частота, которую воспроизвел нервно-мышечный препарат.
2-38. Удельная сила мышцы равна отношению максимального груза к площади физиологического поперечного сечения. В данном случае она равна 8 кг/кв.см. По-видимому, это двуглавая мышца плеча человека.
2-39. В случае, если мембрана повреждена между двумя перехватами Ранвье в мя-котном волокне, возбуждение по волокну будет проходить, так как оно распрост-раняется скачкообразно от перехвата к перехвату. Однако, если энергии потен-циала действия достаточно для возбуждения 2-3 перехватов, то возбуждение будет распространяться и при повреждении волокна в месте одного из перехватов. Это продолжается до тех пор, пока дегенерация не затронет 2-3 межперехватных участка.
2-40. Длина аксона, отделенного от тела клетки, равна 30 см. Полностью она вос-становится только через 100 дней.
2-41. Скорость проведения возбуждения по нервным волокнам типа А - 70-120 мсек. Если расстояние между электродами равно 10 см, то импульс этот путь пройдет за 0,07-0,12 мсек.
2-42. Скорость проведения возбуждения по нервным волокнам типа В - 3-18 м/сек. В данном случае импульс пройдет расстояние между электродами за 4,5-27 мсек.
2-43. Скорость проведения возбуждения по нервам типа С - 0,5-3 м/сек. 5 см. импульс пойдет за 0,1-0,006 сек.
2-44. К волокнам типа С, так как в этом случае скорость проведения возбуж-дения равна 1 м/сек.
2-45. Нерв не возбудится, так как в это время в результате суммации положительных следовых потенциалов мембрана на-ходится в состоянии гиперполяризации, что сопровождается снижением возбуди-мости. Это явление называется посттетаническим торможением (торможением вслед за возбуждением).
2-46. Если между электродами 15 перехватов Ранвье, значит между ними 14 участков. Через каждый из них возбужде-ние перескакивает за 0,07 мсек. Значит, латентный период в данном случае будет равен 0.9 мсек. 2-47. Так как время перескока возбуждения через один межперехватный участок равен 0,07 мсек., то в данном случае таких участков 140: 0,07 = 2000, а перехватов - на один больше, т.е. 2001.
2-48. Так как время движения возбуждения через синапс (синаптическая задержка) равно приблизительно 0,2 мсек, то центральное время рефлекса в данном случае 15*0,2=7,5 мсек.
2-49. Если время синаптической задержки принять за 0,2 мсек, то у центре данного рефлекса находится 500 синапсов.
2-50. Параметрами возбудимости ткани являются: реобаза, полезное время (хро-наксия), лабильность, критический наклон.
2-51. В том и другом случае гальванометр покажет величину, равную потенциалу покоя, так как он одинаков в любом участке клетки.
2-52. Потенциал покоя возникает за счет диффузии ионов калия из клетки в межклеточное пространство. Если бы мембрана была непроницаема для ионов, в том числе и для калия, то ПП был бы равен нулю.
2-53. Так как в этот случае натриевый ток не будет снижать калиевый ток, то величина ПП увеличится.
2-54. ПП уменьшится.
2-55. При выравнивании концентрации натрия по обе стороны мембраны поток этих ионов в клетку при раздражении будет отсутствовать, и ПД не возникнет.
2-56. Инактивация натриевых каналов полностью прекращает процесс деполяризации мембраны и он сменяется реполяризацией, что приводит к восстановлению исходного уровня МП. Если инактивация замедляется, то будет затягиваться аза деполяризации и это вызовет удлинение ПД.
2-57. В нерве и мышце разница между мембранным потенциалом и критическим уровнем деполяризации отличается: в нерве она меньше (20 мв.) в мышце - больше (40 мв.).
2-58. При гиперполяризации возрастает разница между мембранным потенциалом и критическим уровнем деполяризации. При этом для того, чтобы возникло возбуждение, необходима большая сила раздражения.
2-59. Цианиды блокируют работу дыхательных ферментов, прекращают окислительные процессы, которые необходимы для ресинтеза АТФ. При этом в нерве перестанет работать калий- натриевый насос, который поддерживает градиент концентрации ионов по обе стороны мембраны. Концентрация их сравняется, и нерв утратит возбудимость.
2-60. Нерв не может возбуждаться со сколь угодно большой частотой. Этому препятствует абсолютный рефрактерный период, который продолжается примерно 2 мс после начала ПД. При частоте 10 гц интервал между раздражителями составляет 0,1 с, при 100 гц - 0,01 с. В обеих случаях он достаточно велик для того, чтобы рефрактерный период закончился и нерв воспроизвел раздражение с подаваемой частотой. При частоте же 1000 гц интервал между раздражениями слишком мал (0,001 с = 1 мс), и потому каждый второй импульс будет попадать в период рефрактерности после предыдущего возбуждения. Общее число ПД не будет превышать 500.
2-61. ПД уменьшится или вообще не возникнет.
2-62. Если вещество может блокировать ионные каналы или повредить структурные компоненты мембраны, действуя снаружи, то состояние клетки изменится.
2-63. Протеолитические ферменты расщепляют белковые молекулы, которые входят в стенки ионных каналов и с состав «задвижек», открывающих и закрывающих эти каналы. Следовательно, нарушится проницаемость мембраны для ионов. 2-64. Электротравму получит первый человек. Во втором случае величина тока при каждом его колебании нарастает очень быстро, но само колебание продолжается столь малое время, что за него ионы не успевают пройти через мембрану и вызвать деполяризацию. Возбуждение не возникнет. В первом же случае и продолжительность каждого колебания, и скорость нарастания тока достаточны, чтобы вызвать возбуждение. Поэтому сетевой ток напряжением 110 и 220 в и частотой 50 гц опасен для жизни даже при кратковременном воздействии.
2-65. Из-за кратковременности каждого колебания тока ионы не успевают пройти через мембрану и вызвать деполяризацию. Однако при каждой перемене направления тока ионы смещаются от исходного положения. Эти движения частиц приводят к выделению тепловой энергии. Если энергия высокочастотного поля велика, то выделяется много тепла, и может произойти тепловое повреждение ткани.
2-66. В невозбужденном участке нормальная возбудимость, а в том, который был только что возбужден, возникает рефрактерность. Поэтому возбуждение не может вернуться обратно. 2-67. Иннервация обеспечивает не только сократительную деятельность, но и трофические влияния. При денервации нарушается синтез белков в мышце и уменьшается ее масса - происходит атрофия.
2-68. Чтобы ответить на вопрос, надо перейти к прямому раздражению мышцы. Если при этом амплитуда сокращений возрастет, утомление возникло не в мышце, а в синапсах.
2-69. Н-зона - это центральный участок толстой протофибриллы, который не перекрыт актиновыми нитями. При растяжении мышцы степень перекрытия миозиновых нитей уменьшается, так как актиновые нити частично выходят из промежутков между миозиновыми. Соответственно ширина Н-зоны увеличивается.
2-70. При быстром сокращении мостики совершают больше гребковых движений в единицу времени и на это затрачивается больше энергии АТФ.
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-08-26; просмотров: 1142; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.148.144.139 (0.015 с.) |