
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Основные свойства автоволн в АС.Содержание книги
Поиск на нашем сайте
1. Автоволна распространяется без затухания. 2. Автоволны не интерферируют и не отражаются от препятствий. 3. Направление распространения автоволны определяется зонами рефрактерности и покоя. Длина волны возбуждения
Отсюда следует, что если рефрактерность клеток некоторого участка (рис. 6.3,6) повышена по сравнению с R1 на рис. 6.3, а (то есть длительность потенциала действия больше), то и длина волны возбуждения в этом участке больше: В однородных средах, в которых R и V одинаковы в любом участке, длина волны возбуждения постоянна. В таких средах две встречные волны гасят друг друга, поскольку каждая из волн накладывается на невозбудимую зону встречной волны (рис.6.4).
Рис. 6.4. Аннигиляция плоских автоволн в АС Аналогично два встречных фронта пламени степного пожара гасят друг друга. Позади огненного фронта каждого остается черная, выжженная зона - зона рефрактерности, лишенная источников энергии. В неоднородных средах процесс распространения автоволн усложняется. Неоднородной называется активная среда, в различных участках которой значения R и V могут быть не одинаковыми. Активная среда организма, например мышечная ткань, неоднородна. В разных участках мышцы могут проходить кровеносные сосуды, нервные волокна и другие включения. При патологиях, например при возникновении зон некроза, свойства этих зон могут существенно отличаться и по рефрактерности R, и по скорости проведения волны V от этих параметров в участках нормальной мышцы. Очевидно (рис. 6.3), что длины автоволн в различных участках неоднородных активных сред будут неодинаковыми. При выполнении определенных условий это может приводить к сердечным аритмиям.
8. Строение мышцы. Биофизика мышечного сокращения. Мышечная активность - это одно из общих свойств высокоорганизованных живых организмов. Независимо от назначения, особенностей строения и способов регуляции принцип работы различных мышц организма одинаков. Мышечная клетка отличается от других возбудимых клеток свойством,сократимости то есть способность генерировать механическое напряжение и укорачиваться. Мышцы являются генератором тепла, причем не только при мышечной работе, холодовой дрожи, но и в режиме нетонического термогенеза. Мышечная активность в процессе жизнедеятельности обеспечивает работы отдельных органов и целых систем: работу опорно-двигательного аппарата, легких, сосудистую активность, желудочно-кишечного тракта, сократительную способность сердца. Нарушение работы мышц (например, определяющих функционирование легких, сердца) может приводить к патологиям, а ее прекращение - даже к летальному исходу. Мышечная ткань представляет собой совокупность мышечных клеток {волокон), внеклеточного вещества (коллаген, эластин и др.) и густой сети нервных волокон и кровеносных сосудов. Мышцы по строению делятся на: гладкие - мышцы кишечника, стенки сосудов, и поперечно-полосатые - скелетные, мышцы сердца. Независимо от строения все они имеют близкие механические свойства, одинаковый механизм активации и близкий химический состав. Поперечно-полосатая структура мышечных волокон имеет диаметр 20 - 80 мкм и окружено плазматической мембраной толщиной 10 нм. Каждое отдельное волокно -это сильно вытянутая клетка. Длина отдельных волокон (клеток) может существенно варьироваться, в зависимости от вида мышцы, от сотен микрон до нескольких сантиметров. Внутри волокна, кроме известных органелл (ядро, ядрышко, митохондрии, аппарат Гольджи и др.), находятся сократительный аппарат клетки, состоящий из 1000 - 2000 параллельно расположенных миофибрилл диаметром 1-2 мкм, а также клеточные органеллы: саркоплазматический ретикулум и система поперечных трубочек - Т-система.
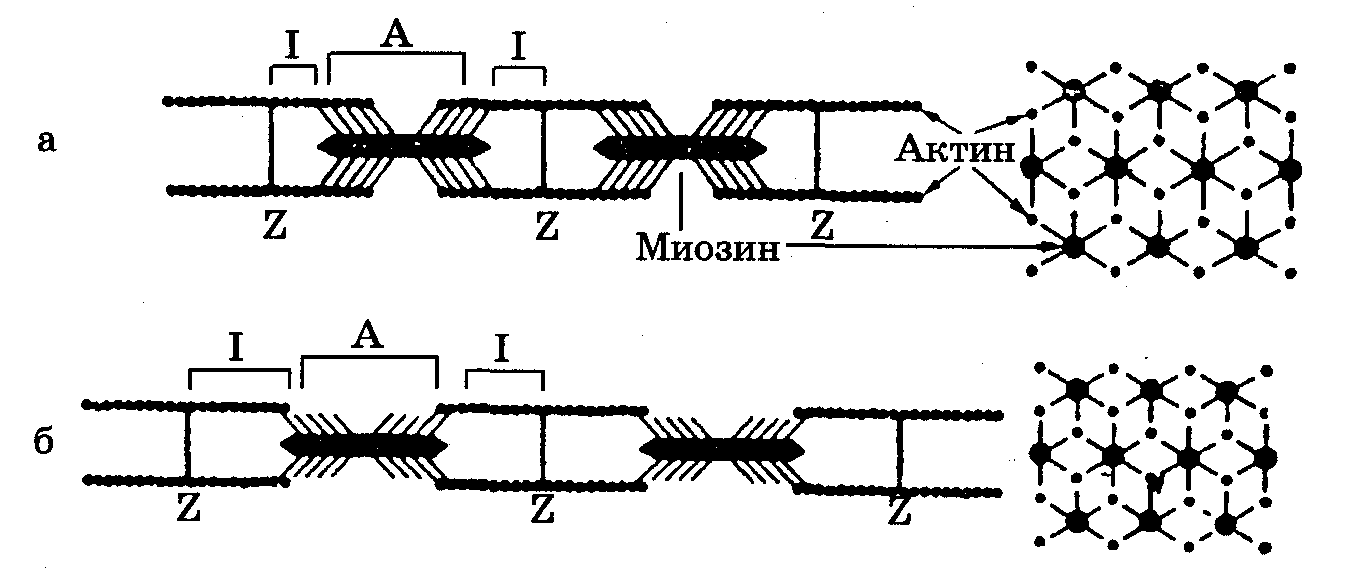
Рис. 7.1. Схематическое изображение миофибриллы мышечного волокна: а – состояние покоя, б - растяжение (подробности в тексте). Справа - схема расположения актина и миозина на поперечном срезе.
В миофибриллах различают (рис. 7.1): А-зону - темные полосы, которые в поляризованном свете дают двойное лучепреломление, то есть обладают свойством анизотропии (отсюда и название: А-зона), 1-зону - светлые полосы, не дающие двойного лучепреломления, то есть изотропные (отсюда название: 1-зона). В области I-зоны проходит темная узкая полоса - Z-диск (от нем. zwischenscheibe - промежуточный диск). Промежуток между двумя Z-дисками называется саркомером и является элементарной сократительной единицей мышечной клетки. Саркомер - это упорядоченная система толстых и тонких нитей, расположенных гексагонально в поперечном сечении. Толстая нить имеет толщину ==12 нм и длину =1,5 мкм и состоит из белка миозина. Тонкая нить имеет диаметр 8 нм, длину 1 мкм и состоит из белка актина, прикрепленного одним концом к Z-диску.
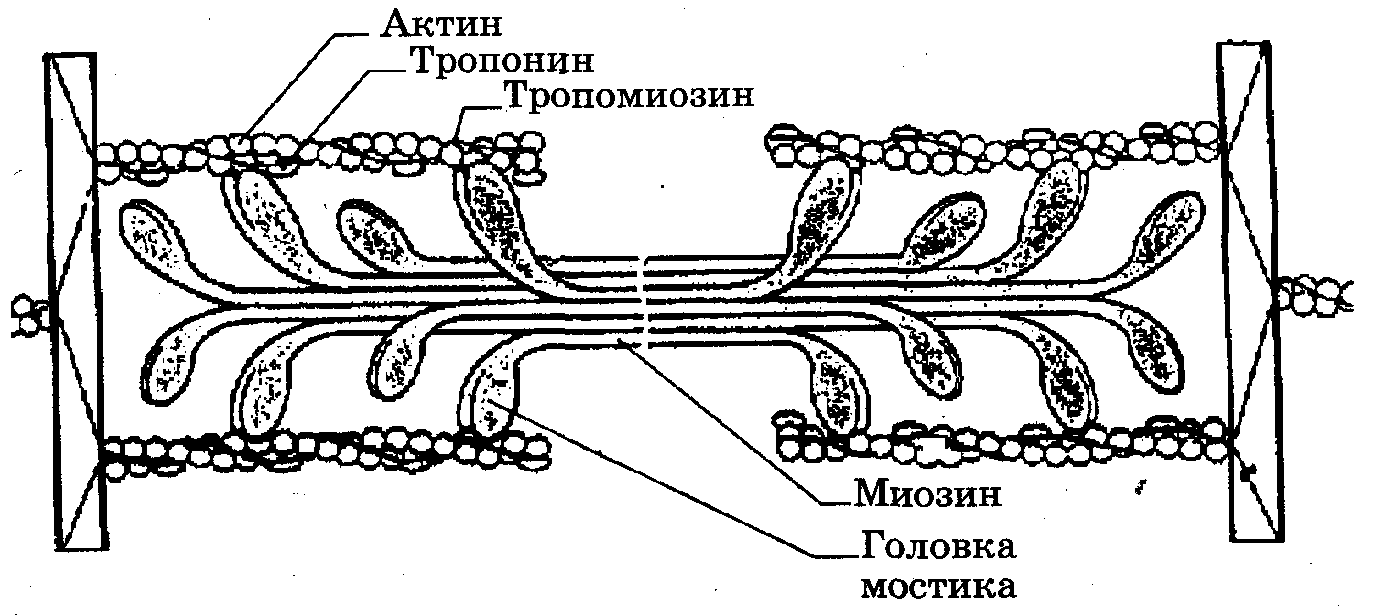
Рис. 7.2. Микроструктура саркомера
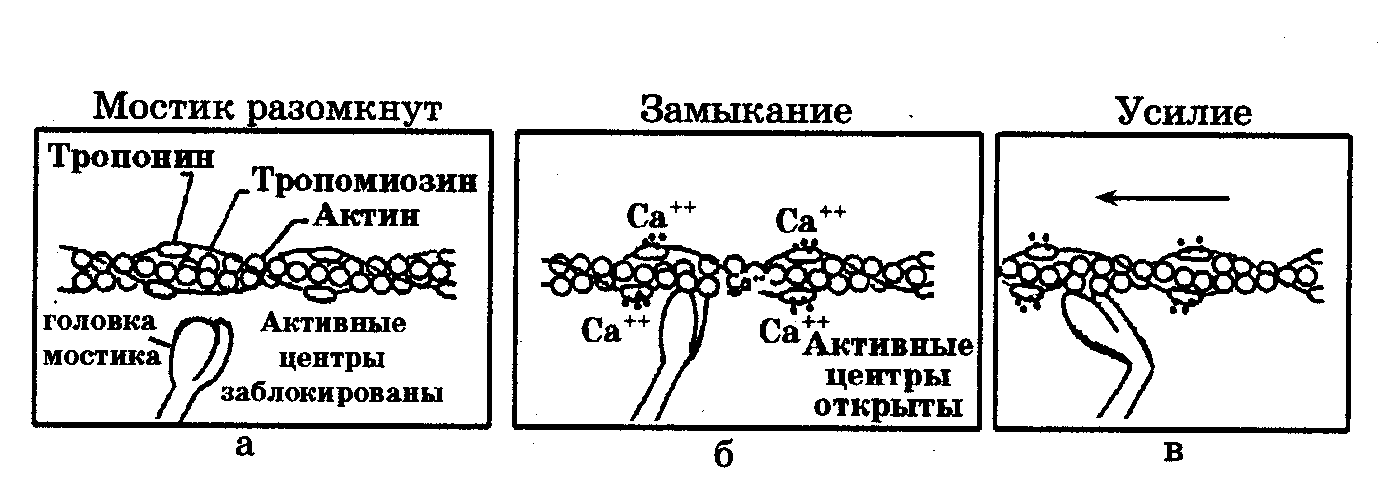
Актиновая нить состоит из двух закрученных один вокруг другого мономеров актина толщиной по 5 нм (рис. 7.2). Эта структура похожа на две нитки бус, скрученные по 14 бусин в витке. В цепях актина регулярно примерно через 40 нм встроены молекулы тропонина, а сама цепь охватывает нить тропомиозина. При сокращении мышцы тонкие нити вдвигаются между толстыми. Происходит относительное скольжение нитей без изменения их длины. Этот процесс обусловлен взаимодействием особых выступов миозина - поперечных мостиков с активными центрами, расположенными на актине. Мостики отходят от толстой нити периодично на расстоянии 14,5 нм друг от друга. В расслабленном состоянии миофибрилл молекулы тропомиозина блокируют прикрепление поперечных мостиков к актиновым цепям (рис. 7.3, а).
Рис. 7.3. Процесс активации мостика и генерации усилия в саркомере
Ионы Са2+ активируют мостики и открывают участки их прикрепления к актину (рис. 7.3,6). В результате мостики миозина прикрепляются к актиновым нитям, расщепляются молекулы АТФ и изменяется конформация мостиков: их головки поворачиваются внутрь саркомера (рис. 7.3, в). Это приводит к генерации силы, скольжению актина относительно толстой нити миозина к центру саркомера, что вызывает укорочение мышцы. После окончания активации мостик размыкается и саркомер возвращается в исходное состояние. При укорочении объем саркомера практически не меняется, а следовательно, он становится толще, что и подтверждается на снимках поперечного сечения мышц с помощью электронной микроскопии. Каждый цикл замыкание-размыкание сопровождается расщеплением одной молекулы АТФ. Таким образом, актин-миозиновый комплекс является механохимическим преобразователем энергии АТФ. Рассмотренная структура и последовательность процессов называется моделью скользящих нитей. Впервые скольжение нитей в саркомере было обнаружено английским ученым Х.Хаксли. Он же сформулировал модель скользящих нитей. Существенный вклад в разработку теории скользящих нитей внес В.И. Дещеревский. Представленная структурная модель скользящих нитей надежно подтверждена экспериментально и является опытным фактом, на котором основаны все современные теории мышечного сокращения. Экспериментальные данные о микроструктуре мышц были изучены с помощью электронной микроскопии, рентгеноструктурного анализа и метода дифракции синхротронного излучения. Основные положения модели скользящих нитей: 1. Длины нитей актина и миозина в ходе сокращения не меняются. 2. Изменение длины саркомера при сокращении - результат относительного продольного смещения нитей актина и миозина. 3. Поперечные мостики, отходящие от миозина, могут присоединяться к комплементарным центрам актина. 4. Мостики прикрепляются к актину не одновременно. 5. Замкнувшиеся мостики подвергаются структурному переходу, при котором они развивают усилие, после чего происходит их размыкание. 6. Сокращение и расслабление мышцы состоит в нарастании и последующем уменьшении числа мостиков, совершающих цикл замыкание-размыкание. 7. Каждый цикл связан с гидролизом одной молекулы АТФ. 8. Акты замыкания-размыкания мостиков происходят не зависимо друг от друга. Мышцы можно представить как сплошную среду, то есть среду, состоящую из большого числа элементов, взаимодействующих между собой без соударений и находящихся в поле внешних сил. Мышца одновременно обладает свойством упругости и вязкости, то есть является вязко-упругой средой. Для такой среды предполагаются справедливыми законы классической механики. Активное сокращение мышцы. Для исследования характеристик сокращающихся мышц используют два искусственных режима: 1. Изометрический режим, при котором длина мышцы I — const, а регистрируется развиваемая сила F(t). 2. Изотонический режим, при котором мышца поднимает постоянный груз Р — const, а регистрируется изменение ее длины во времени Al (t).
Зависимость скорости укорочения от нагрузки Р является важнейшей при изучении работы мышцы, так как позволяет выявить закономерности мышечного сокращения и его энергетики. Она была подробно изучена при разных режимах сокращения Хиллом и представлена на рис. 7.8. Им же было предложено эмпирическое выражение, описывающее эту кривую:
Уравнение Хилла является основным характеристическим уравнением механики мышечного сокращения. Ро - максимальное изометрическое напряжение, развиваемое мышцей, или максимальный груз, удерживаемый мышцей без ее удлинения; b - константа, имеющая размерность скорости, а - константа, имеющая размерность силы.
Эффективность работы мышцы при сокращении может быть определена как отношение совершенной работы к затраченной энергии АЕ: Развитие наибольшей мощности и эффективности сокращения достигается при усилиях 0,3 - 0,4 от максимальной изометрической нагрузки Ро для данной мышцы. Это используют, например, спортсмены-велогонщики: при переходе с равнины на горный участок нагрузка на мышцы возрастает и спортсмен переключает скорость на низшую передачу, тем самым уменьшая Р, приближая ее к Ропт. Кинетическая модель Дещеревского позволила объяснить феноменологическое уравнение Хилла, смысл констант а и Ь, входящих в него, смысл величины максимальной силы Ро изометрического сокращения. Кроме того, показано, что внутренняя вязкая компонента мышцы определяется силой, развиваемой тормозящими мостиками, число которых растет с увеличением скорости сокращения мышцы. Электромеханическое сопряжение - это цикл последовательных процессов, начинающийся с возникновения потенциала действия ПД на сарколемме (клеточной мембране) и заканчивающийся сократительным ответом мышцы. Нарушение последовательности процессов сопряжения может приводить к патологиям и даже к летальному исходу. Процесс сокращения кардиомиоцита происходит следующим образом В кардиомиоците электромеханическое сопряжение идет в две ступени: вначале небольшой входящий поток кальция активирует мембраны СР, способствуя большему выбросу кальция из внутриклеточного депо, а затем в результате этого выброса происходит сокращение саркомера. Описанный выше двухступенчатый процесс сопряжения доказан экспериментально. Опыты показали, что: а) отсутствие потока кальция извне клетки jCa прекращает сокращение саркомеров, б) в условиях постоянства количества кальция, высвобождаемого из СР, изменение амплитуды потока кальция приводит к хорошо коррелирующему изменению силы сокращения. Поток ионов Са2+ внутрь клетки выполняет таким образом две функции: формирует длительное (200 мс) плато потенциала действия кардиомиоцита и участвует в процессе электромеханического сопряжения. Следует отметить, что не во всех мышечных клетках организма процесс сопряжения происходит, как в кардиомиоците. Так, в скелетных мышцах теплокровных потенциал действия короткий (2-3 мс) и медленный поток ионов кальция в них отсутствует. В этих клетках сильно развита Т-система поперечных трубочек, подходящих непосредственно к саркомерам близко к z-дискам (см. рис. 7.11). Изменения мембранного потенциала во время деполяризации через Т-систему передается в таких клетках непосредственно на мембрану СР, вызывая залповое высвобождение ионов Са2+ и дальнейшую активацию сокращения (3, 4, 5). Временной ход описанных процессов показан на рис. 7.12.
Рис. 7.12. Временное соотношение между потенциалом действия кардиомиоцита (а) и одиночным сокращением (б) в этих клетках. Ордината слева - мембранный потенциал, справа - сила.
Общим для любых мышечных клеток является процесс освобождения ионов Са2+ и внутриклеточных депо - саркоплазматического ретикулума и дальнейшая активация сокращения. Задержка начала развития сокращения в скелетных мышцах составляет 20 мс, а в сердечной - несколько больше (до 100 мс).
9. Принцип автоматической регуляции в живых системах.
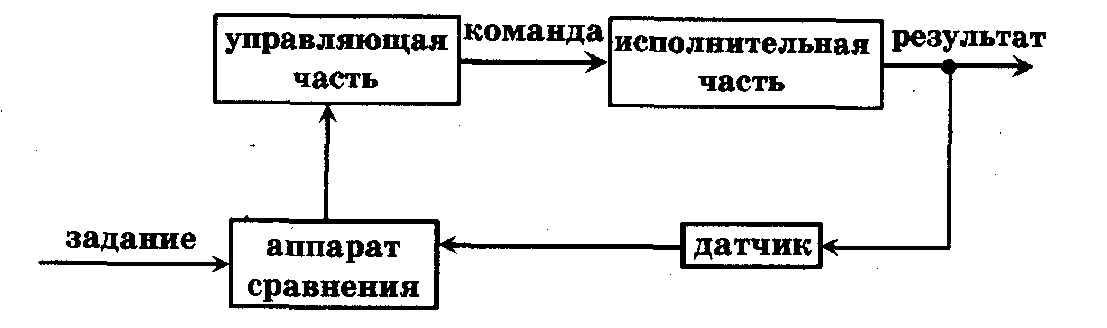
Биологическая кибернетика является составной частью биофизики сложных систем. Биологическая кибернетика имеет большое значение для развития современной биологии, медицины и экологии. Кибернетический подход к изучению функционирования органов, физиологических систем, организма в целом и сообществ организмов позволяет изучить особенности управления и саморегуляции биологических объектов в норме и патологии. Управление (регулирование) - процесс изменения состояния или режима функционирования системы в соответствии с поставленной перед ней задачей. Всякая система содержит управляющую часть и исполнительную часть (рис. 10.2). По линии прямой связи управляющая часть посылает в соответствии с заданием в исполнительную часть команды. По линии обратной связи в управляющую часть поступают сведения из исполнительной части о выполнении команд. Например, головной мозг (управляющая часть) посылает команды мышцам, а по каналам обратной связи в головной мозг поступают сведения о выполнении команды (например, о соответствующем двигательном акте).
Рис. 10.2. Простейшая схема системы управления
Биологические объекты относятся к саморегулируемым системам. Саморегулируемые системы такие, которые обладают способностью поддерживать свое состояние или режим функционирования на определенном заданном уровне при непредвиденных внешних воздействиях.
Рис. 10.3. Система регулирования по возмущению
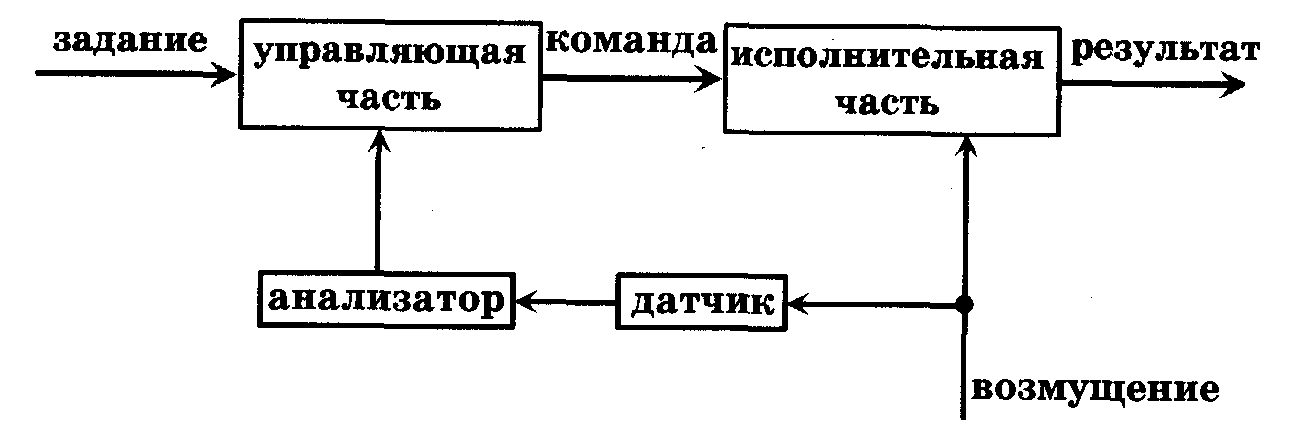
Теория автоматического регулирования выделяет два основных способа регулирования: регулирование по возмущению и регулирование по отклонению. Система регулирования (рис. 10.3) по возмущению позволяет устранить результаты непредвиденного внешнего воздействия на систему с целью сохранить заданный режим функционирования. Для этого система должна содержать в своей памяти информацию о возможных последствиях возмущения. Примером регулирования по возмущению является система терморегуляции организма, основанная на сигналах кожных терморецепторов об изменении температуры окружающего воздуха. Другим распространенным видом регулирования является регулирование по отклонению. В случае регулирования по отклонению (рис. 10.4) управляющая часть вырабатывает команды, вызывающие изменения в системе, компенсирующие отклонения от заданного режима функционирования в системе.
Рис. 10.4. Система регулирования по отклонению
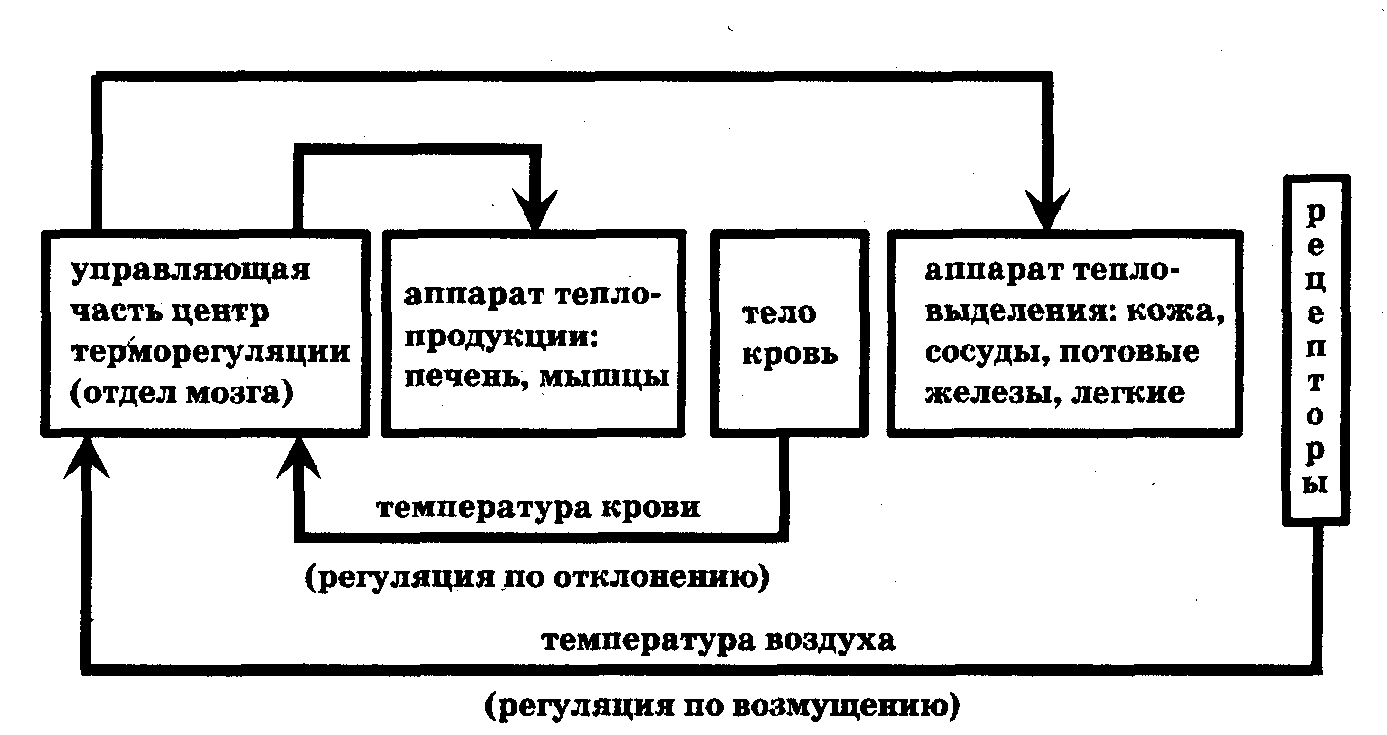
Датчик передает по каналу обратной связи сведения о режиме функционирования системы в аппарат сравнения, в котором они сравниваются с заданными параметрами, в случае отклонения от задания (рассогласования) управляющая часть вырабатывает команды, устраняющие отклонения. Обратные связи - необходимое условие процессов саморегуляции. Обратная связь передает информацию о результате функционирования системы в управляющую часть. Различают положительные и отрицательные обратные связи. Положительные обратные связи приводят к выработке команд, ведущих к увеличению отклонения системы от первоначального состояния. Например, всасывание желудком продуктов переваривания белков приводит к увеличению сокоотделения («аппетит приходит во время еды»). В то же время некоторые патологии развиваются с участием положительных обратных связей. Например, ослабление сердечной деятельности может приводить к закупорке сосудов, что в свою очередь вызывает еще большее ослабление сердечной деятельности. Отрицательные обратные связи вызывают команды, стремящиеся уменьшить отклонения в системе. Регулирование по отклонению осуществляется с использованием отрицательных обратных связей. Например, при перегреве организма усиливается потоотделение, учащается дыхание, что приводит к увеличению теплоотдачи в окружающую среду и понижению температуры организма. Биологические системы обладают способностью менять задание, программу, определяющую результат их функционирования, в целях улучшения результата деятельности или приспособления к резким изменениям условий внешней среды. Механизм приспособления организма к меняющимся условиям называется гомеостазом. Гомеостаз обеспечивает поддержание в организме условий постоянного функционирования процессов метаболизма: постоянство температуры, рН, давления и т.д. Более строго, параметры во внутренней среде организма непостоянны. Они совершают колебания около средних значений: сезонные, суточные, за цикл дыхания, сердечного сокращения и т.д. Поддержание колебаний параметров системы на постоянном уровне (по амплитуде и частоте) называется гомеокинезом. Рассмотрим в качестве примера упрощенную схему терморегуляции теплокровных организмов (рис. 10.5).
Рис. 10.5. Схема терморегуляции теплокровных организмов
Информация о температуре тела организма поступает в центр терморегуляции головного мозга вместе с омывающей его кровью. При отклонении от заданной температуры вырабатываются команды в исполняющую часть для компенсации отклонения. Это регуляция по отклонению при помощи отрицательной обратной связи. Кроме того, осуществляется регуляция по возмущению на основе сведений об изменении температуры окружающего воздуха, поступающих от кожных терморецепторов. Команды из центра терморегуляции поступают в аппарат теплопродукции (в основном тепло вырабатывает печень, мышцы) и в аппарат тепловыделения (кожа, сосуды, потовые железы, легкие). Так, при перегреве организма уменьшается теплообразование и усиливаются процессы тепловыделения в основном посредством испарения воды с поверхности кожи и легких.
10. Влияние изменения радиоактивного фона Земли на состояние человека.
Естественный радиоактивный фон и фон электромагнитных излучений являются сферой нашего обитания, неотъемлемой частью и условием развития жизни. Космические события, изменения солнечной активности, нарушение магнитосферы и ионосферы Земли могут оказывать влияние на жизненные процессы всех биологических объектов нашей планеты. На биосферу Земли непрерывно действует космическое излучение, а также потоки альфа- и бета-частиц, гамма-квантов в результате излучения различных радионуклидов, рассеянных в земной коре, воде подземных источников, реках, морях и океанах, в воздухе. Кроме того, радионуклиды входят в состав живых организмов. Совокупность излучений этих радиоактивных источников называется природным или естественным радиоактивным фоном. Радиационный фон Земли определяется в основном следующими природными источниками: радионуклиды - это 220Rn и 222Rn (50%), 40К (15%), а также радионуклиды, составляющие ряды урана (20%) и космические лучи (15%). Изотопа 40К содержится в почве, удобрениях, а также в головном мозге, мышцах, селезенке и костном мозге. Период полураспада 40К составляет 1,3 • 109 лет Космические лучи возникают в результате межзвездных и галактических событий и активности Солнца. Космическое излучение состоит из потоков протонов высоких энергий, альфа-частиц, ядер некоторых элементов, потоков электронов, фотонов и нейтронов. Магнитное поле Земли отклоняет низкоэнергетические заряженные частицы. Частицы высоких энергий, взаимодействуя с атмосферой, образуют в результате ядерных реакций целую серию радионуклидов 3Н, 7Ве, 22Na и др. и потоки нейтронов и протонов. Образуются космические ливни, составляющие вторичное космическое излучение, проникающее в нижние слои атмосферы. На биосферу воздействует ионизирующий компонент вторичного космического излучения. Оно дает 1,9-2,5 ионизации / см3 за 1 с на уровне моря; в горах в 2-3 раза выше. Диапазон эквивалентных доз для человека от природных источников, создающих радиоактивный фон, мкЗв за год:
внешнее облучение; внутреннее облучение; космические лучи - 300 40К - 180 40К -120 222Rn - 800 ряды 238U и 232Th - 230 210Rn - 130.
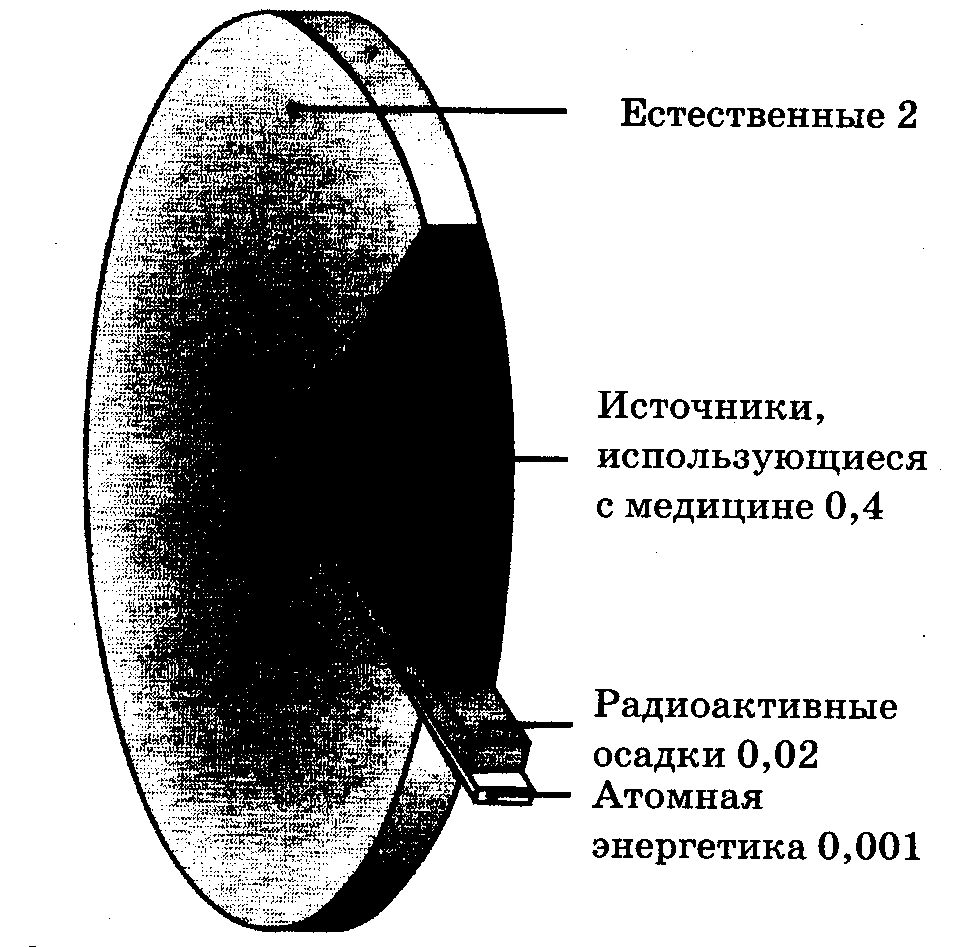
Средняя мощность дозы облучения от всех источников природной радиации на гонады, гаверсовы костные каналы и костный мозг человека как наиболее чувствительные ткани к действию радиации составляет около 200 мбэр в год (2 мЗв в год). Однако радиационный фон в зависимости от местоположения, времени года, наличия промышленных предприятий и др. может меняться в значительных пределах. Так, в городах мощность дозы фонового излучения составляет в среднем около 20 мкР/час и может меняться в 2 и более раза в различных районах города. На рис. 11.4 представлены источники радиации и дозы естественного фона и фона, вызванного активной деятельностью человека при нормальных условиях функционирования.
Цифры указывают величину дозы в миллизевертах
Рис. 11.4. Средние годовые эффективные эквивалентные дозы облучения от естественных и техногенных источников радиации
Природный радиоактивный фон, оказывая влияние на развитие жизни на Земле, является неотъемлемой частью сферы обитания человека. В условиях радиоактивного фона происходят такие процессы, как деление одноклеточных организмов и клеток, развитие эмбрионов насекомых, рост и развитие высших растений и животных. Нарушения радиоактивного фона в локальных условиях и тем более глобальные опасны для существования биосферы и могут привести к непоправимым последствиям. Причиной увеличения радиоактивного фона является активная деятельность человека. Создание крупной промышленности, научных установок, энергетических источников, военной техники и др. может приводить к локальным изменениям фона. Но наиболее опасными причинами нарушений естественного радиоактивного фона являются выбросы радиоактивных частиц, которые могут возникнуть при ядерных взрывах или при эксплуатации атомных электростанций (АЭС). Разряженный нагретый воздух поднимает вверх продукты распада на высоту до 50 км, после чего это облако может расплываться на сотни и даже тысячи километров. Радиоактивные частицы выпадают на поверхность земли, образуя радиоактивный след. Радионуклиды, находящиеся в виде аэрозолей в воздухе, а также осевшие на земную поверхность, могут представлять для человека опасность. Оценку степени опасности можно получить по активности препарата А:
где N - количество распадающихся ядер. Активность данного препарата измеряется в кюри (Ки): 1 Ки = 3,7 • 1010 распад/с. Активность уменьшается со временем по экспоненциальному закону:
где
где r - расстояние от источника излучения, К - гамма-постоянная, зависящая от природы радиоактивного источника. Значения 131I -2,6 60Со -13,5 24Na -18,6 22ORa -8,4 Таким образом, при выпадении радионуклидов на почву степень опасности их влияния на организм зависит от природы радиоактивного изотопа (
где r - расстояние от источника излучения, К - гамма-постоянная, зависящая от природы радиоактивного источника. А - активность препарата, Таблица 11.4.
|
||||
|
|
Последнее изменение этой страницы: 2016-08-25; просмотров: 705; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.118.10.75 (0.013 с.) |

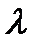 , определяется соотношением, введенным Винером:
, определяется соотношением, введенным Винером: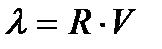 (6.4)
(6.4)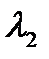 >
> 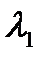 . Длины волн возбуждения для различных отделов сердца указаны в § 17.
. Длины волн возбуждения для различных отделов сердца указаны в § 17.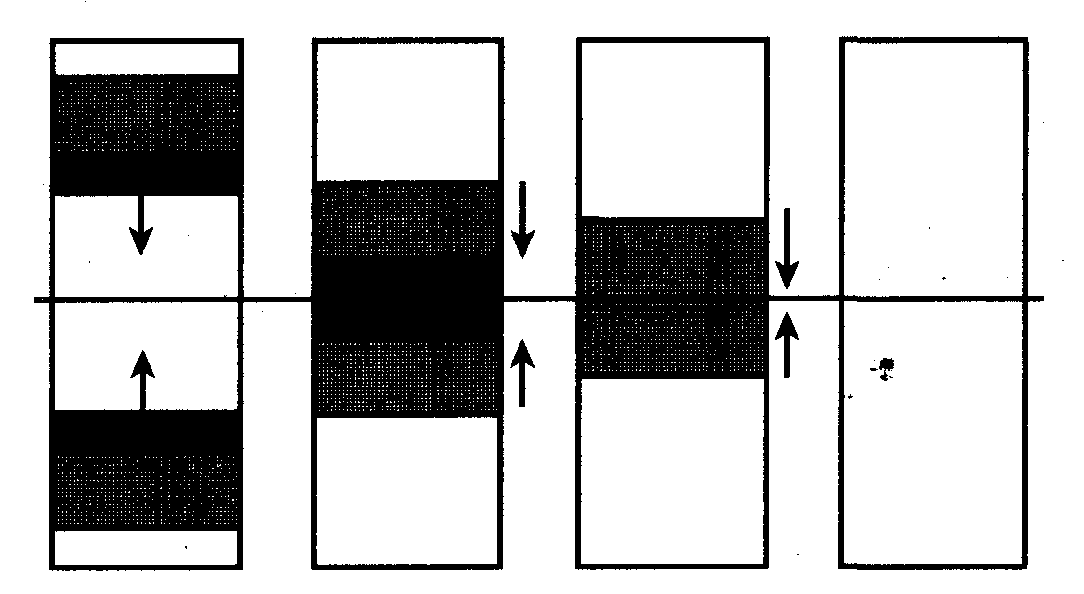
 . (7.2)
. (7.2)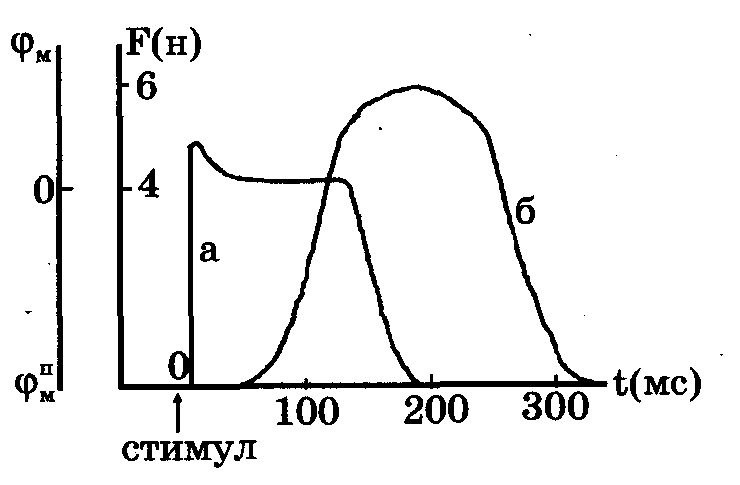
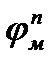 - потенциал покоя
- потенциал покоя
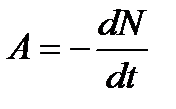 ,
,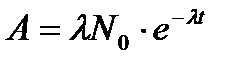 , (11.13)
, (11.13) - постоянная распада, N0 - начальное количество ядер. Для точечных источников излучений мощность экспозиционной дозы уменьшается с расстоянием по закону:
- постоянная распада, N0 - начальное количество ядер. Для точечных источников излучений мощность экспозиционной дозы уменьшается с расстоянием по закону: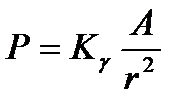 , (11.14)
, (11.14) для некоторых радиоактивных источников:
для некоторых радиоактивных источников: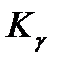 ), его активности и расстояния г от человека до источника, а экспозиционную дозу можно оценить из соотношения
), его активности и расстояния г от человека до источника, а экспозиционную дозу можно оценить из соотношения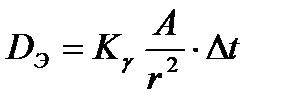 ,
, t - время облучения.
t - время облучения.


