Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Сплайсинг – способ создания многообразия белковСодержание книги
Поиск на нашем сайте
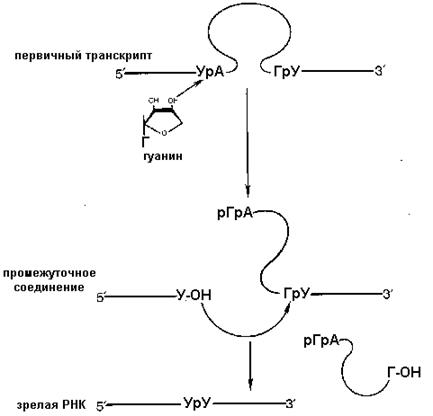
После модификации концов иРНК наступает одна из самых сложных реакций процессинга -точное вырезание различных по длине внутренних участков (интронов) и сшивание оставшихся, несущих смысловую нагрузку для кодируемого белка - экзонов..Механизм сплайсинга интронов группы I.
ность реакций, происходяших при этом получила название сплайсинг. Анализ большого количества генов иРНК позволил идентифицировать высоко консервативные консенсусные последовательности на 5 ' и 3 ' концах практически всех интронов иРНК. 3 ' конец 5 ' экзона называют донорским участком сплайсинга, а 5 ' конец 3 ' экзона - акцепторным участком сплайсинга Существуют четыре различных класса интронов. 2 наиболее распространенных – объединяют в группу I и группу II интронов. Группа I интронов найдена в ядерных, митохондриальных и хлоропластных генах р-РНК, группа II в митохондриальных и хлоропластных генах и- РНК. Большинство интронов из группы I и группы II подвергаются автосплайсингу, то есть для их сплайсинга не требуется дополнительных белковых факторов. Роль катализатора этого процесса выполняет сама РНК.
Последовательность нуклеотидов в зоне контакта интронов и экзонов.
Интроны группы I используют внешний гуанозиловый нуклеотид в качестве кофактора. 3 ' – ОН этого нуклеотида действует как нуклеофил, атакуя 5 ' - фосфат 5 ' нуклеотида интрона.
Механизм сплайсинга интронов группы II.
Образующаяся 3 '–ОН группа 3' конца 5' экзона затем атакует 5 ' нуклеотид 3 ' экзона, высвобождая интрон, а донорный участок сплайсинга соединяется с акцепторным участком другого экзона ковалентным типом связи Интроны группы II подвергаются сплайсингу практически таким же способом одна
Интроны группы II подвергаются сплайсингу практически таким же способом однако роль нуклеофила выполняет 2'-ОН группа остатка аденина самого интрона (рис.9-43). Этот остаток атакует 3 ' нуклеотид 5 ' экзона, формируя внутреннюю петлю называемую структурой аркана или лассо. 3 ' конец 5 ' экзона затем атакует 5 ' конец 3 ' экзона как и при сплайсине в группе I, высвобождая интрон и ковалентно соединяя два экзона вместе Третий класс интронов – еще один большой класс интронов, найденный в ядерных иРНК. Этот класс интронов подвергается реакции сплайсинга, подобно группе II интронов при котором образуется внутренняя структура аркана. Однако, сплайсинг катализируется специализированными комплексами РНК-белок, которые названы маленькими ядерными рибонуклеопротеидными частицами (snRNPs). РНК, найденные в snRNPs идентифицированы как U1, U2, U4, U5 и U6. Гены, кодирующие эти snRNAs высоко консервативны у позвоночных и насекомых, они также найдены в дрожжах и у улиток, что указывает на их важность. U1 РНК имеет последовательности, которые являются комплементарными последовательностям вблизи 5 ' конца интрона. Связывание U1 РНК отличает ГУ на 5 ' конце интрона от других беспорядочно размещенных ГУ последовательностей в иРНК. U2 РНК также узнает последовательности в интроне, в этом случае вблизи 3 ' конца. Присоединение U4, U5 и U6 РНК формирует комплекс, идентифицированный как сплайсеосома, который удаляет интрон и соединяет два экзона вместе. Четвертый класс интронов найден в некоторых т-РНК. Сплайсинг интронов этого класса катализируется специфической сплайсирующей эндонуклеазой, которая использует энергию гидролиза АТФ, катализируя удаление интрона и соединение экзонов. На первый взгляд присутствие интронов в генах эукариот, казалось бы, является бессмысленной тратой клеточной энергии затрачиваемой вначале на их включение в первичный транскрипт только для того, чтобы быть удаленным позже. Однако, присутствие интронов защищают функционально активную часть генома клетки от повреждающего действия химических илифизических (лучевых) факторов. Еще одна важная функция интронов позволяет при помощи так называемого альтернативного сплайсинга увеличить генетическое разнообразие генома без увеличения числа генов. В результате изменения распределение экзонов одного транскрипта во время сплайсинга возникают различные РНК и следовательно различные белки. Альтернативный сплайсинг происходить или в определенных стадиях развития организма или в различных типах клеток.
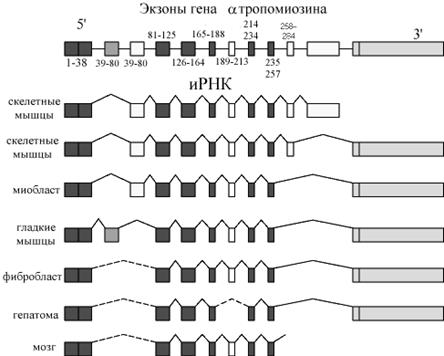
Известны уже более 40 генов, транскрипты которых подвергаются альтернативному сплайсингу. Например, транскрипт гена кальцитонина, в результате альтернативного сплайсинга дает РНК, которая служит матрицей для синтеза кальцитонина (в щитовидной железе) или специфический белок, называемый белок, связанный с геном кальцитонина (CGRP, в мозге). Еще более сложному альтернативному сплайсингу подвергается транскрипт гена a-тропомиозина. Были идентифицированы по крайней мере 8 различных тропомиозиновых иРНК, полученных из одного транскрипта (см рис) Нарушения процессов сплайсинга может вести к различным заболеваниям. Например, b талассемии связаны с дефектами генов, кодирующих b–глобины. Некоторые из этих дефектов вызываются мутациями в последовательностях гена, обеспечивающих узнавание интронов.
Схематическое изображение сплайсинга первичного транскрипта гена a-тропомиозина
В результате происходит нарушение процессинга первичного транскрипта гена b - глобина. Еще один тип заболеваний, связанный с нарушением сплайсинга связан с образованием антител к белкам, участвующим в сплайсинге. Так, например, причиной системной красной волчанки - одного из тяжелых заболеваний соединительной ткани, является образование антител к комплексу белок - U1 РНК сплайсеосомы. РНК-полимераза III транскрибирует гены 5S РНК, тРНК и 7SL РНК, которые кодируют РНК, имеющие самостоятельное функциональное значение. Гены 5S РНК, как и гены других рибосомальных РНК, обычно представлены сотнями или даже тысячами тандемно повторяющихся копий, разделенных спейсерами. Эти гены собраны в одном или нескольких районах генома. Гены разных тРНК (в эукариотической клетке насчитывается 40-60 основных типов тРНК) также часто сгруппированы в кластеры, расположенные в разных хромосомах. Часть гяРНК также транскрибируется РНК-полимеразой III. Клетки высших эукариот содержат около 20000 молекул РНК-полимеразы III, точное количество этого фермента зависит от скорости роста клетки.. Транспортные РНК также синтезируются в форме больших молекул предшественников, которые затем подвергаются процессингу. Один из предшественников тРНК E. coli –представляет первичный транскрипт в 950 нуклеотидов длиной, который расщепляется рибонуклеазой P на 7 различных молекул тРНК. Рибонуклеаза P – рибонуклеопротеид, содержащий молекулу РНК в 377 нуклеотидов, которая является фактическим ферментом. Это был первый пример так называемых рибозимов – ферментов, которые представляют собой РНК, а не полипептидные цепи. После завершения расщепления первичного транскрипта тРНК происходит удаление дополнительных нуклеотидов на 5 ' и 3 ' концах и к 3' концу каждой тРНК присоединяется последовательность 5 '-ЦЦА-3',а несколько нуклеотидов основной структуры подвергаются модификации. В т-РНК были идентифицированы больше 60 различных измененных оснований.
Организационно- Методические указания: 1.Мультимедийная презентация доцент каф. Свергун В.Т. Дата
|
||||||||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 430; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.119 (0.007 с.) |