Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Нарушение обмена пиримидиновых нуклеотидов также приводит к болезням.Содержание книги
Поиск на нашем сайте
Конечными продуктами метаболизма пиримидинов являются хорошо растворимые в воде соединения, такие, как СО2, аммиак, b-аланин и b-изо-пропионат. Поэтому клинические симптомы при их усиленном образовании проявляются слабо. В случаях гиперурикемии, обусловленной избыточной продукцией ФРПФ, наблюдаются повышенный синтез пиримидиновых нуклеотидов и соответственно увеличивается выделение конечных продуктов метаболизма, в частности b-аланина. Поскольку для синтеза тимидилата необходим N5, N'°-метилентетрагидрофолат, нарушения метаболизма фолата и витамина В12; приводят к дефициту ТМР (влияние недостатка витамин В12 реализуется опосредованно). Экскреция с мочой b-аминоизобутирата—аутосомный рецессивный признак. Он распространен главным образом среди жителей Востока Описаны два типа первичной наследственной оро товой ацидурии. Более распространенная форма (хотя и встречающаяся весьма редко) связана с утратой двух ферментов—оротат-фосфорибозилтрансферазы и оротидилат(ОМР)-декарбоксилазы. В детском возрасте для больных характерны отставание в развитии, мегалобластическая анемия и «оранжевая кристаллоурия» (оротовая кислота). При отсутствии лечения пиримидиновыми нуклеозидами пациенты подвержены инфекциям. Второй тип наследуемой оротовой ацидурии (тип 11) связан с недостатком только ОМР-декарбоксилазы.
У пациентов, страдающих оротовой ацидурией первого типа, основным продуктом экскреции является оротовая кислота. У больных aцидурией второго типа с мочой выделяется главным образом оротидин и только небольшое количество оротовой кислоты. В эритроцитах пациентов с оротовой ацидурией типа I были значительно повышены удельные активности аспартаттранскарбамоилазы и дегидрооротазы, однако, прием уридина восстанавливал нарушения. Эти наблюдения свидетельствуют о том, что конечные продукты пути биосинтеза пиримидинов регулируют активность рассматриваемых ферментов. В условиях недостатка конечных продуктов происходит дерепрессия, вероятно координированного характера, по крайней мере этих двух ферментов. У больных с недостаточностью орнитинтранскарбамоилазы — митохондриального фермента клеток печени, ответственного за раннюю стадию синтеза мочевины и аргинина, наблюдается увеличение экcкpeции оротовой кислоты, урацила и уридина. По-видимому, в митохондриях этих пациентов в результате снижения активности орнитинтранскарбамоилазы накапливается карбамоилфосфат, который выходит в цитозоль. где используется как cyбстрат для синтеза пиримидиновых нуклеотидов de novo. При избыточном образовании оротовой кислоты диагностируется оротовая ацилурия. Обычно она проявляется в легкой форме и не сопровождается образованием кристаллов. Синтез и экскреция оротовой кислоты у таких больных усиливаются при употреблении в пищу мясных продуктов таких, как мясо. Оротовую ацидурию может вызвать применение некоторых лекарственных препаратов. Упоминаемый выше аллопуринол, используемый для лечения некоторых форм подагры фосфорибозилируется оротат-фосфорибозилтрансферазой, конкурентно ингибируя фосфорибозилирование оротовой кислоты. Более того, образующийся при этом необычный нуклеотид подавляет оротидилатдекарбоксилазу. вызывая тем самым оротовую ацидурию и оротидинурию. Однако путь биосинтеза пиримидинов у человека, постепенно «приспосабливается» к такому ингибированию и симптомы оротовой ацидурии проявляются только на ранних стадиях лечения препаратом
6-азауридин после превращения в 6-азауридилат функционирует как конкурентный ингибитор ОМР- декарбоксилазы., что приводит к увеличению экскреции оротовой ксилоты и оротидина. При специфическом поражении митохондрий клеток печени (синдром Рейе) имеет место вторичная оротовая ацидурия. Дело в том, что пораженные митохондрии оказываются неспособными утилизировать карбамоилфосфат, который как и в случае наследуемой недостаточности орнитинтранскарбамоилазы, вызывает избыточное образование оротовой кислоты
Биосинтез ДНК – один из важнейших процессов передачи генетической информации последующим поколениям и ее хранения. Некоторые их этих процессов проходят во всех типах клеток (репликация и репарация ДНК, синтез ДНК митохондрий), другие же - только в специализированных (например, осуществляющих синтез антител, транспозонов) или инфицированных вирусами клетках (репликация бактериофагов и вирусов). Все эти процессы катализируются разными ДНК- полимеразами, различающимися по числу субъединиц, молекулярной массе, ассоциацией с разными вспомогательными белками, ускоряющими процессы биосинтеза ДНК, и по многим другим структурным свойствам и функциональному назначению. В целом общие принципы синтеза ДНК схожи, однако, несомненно, существуют и различия, обусловленные видовыми и клеточными особенностями. Как уже упоминалось выше синтез ДНК в подавляющем большинстве случаев проходит по полуконсервативному механизму, то есть дочерняя цепь ДНК синтезируется на материнской цепи, называемой обычно матрицей. Матрицами могут быть односпиральные ДНК или РНК. Ферменты, катализирующие синтез ДНК - полимеразы различаются по многим показателям. Одни из них синтезируют дочерние цепи ДНК процессивно, то есть, диссоциируя из комплекса с матричной цепью (М-дДНК) после удлинения дДНК на десятки нуклеотидных остатков (вплоть до полного окончания процесса), другие же функционируют дистрибютивно, диссоциируя из комплекса с матрицей практически после каждого шага удлинения. Процессивность полимераз усиливается специальными дополнительными беловыми факторами. Все ДНК-полимеразы уждаются для начала своей работы в предварительно синтезированных олигонуклеотидах – затравках, 3’-ОН группа которых используется для удлинения цепи. (Эти короткие отрезки, обычно, рибонуклеотидов синтезируются специальными РНК-полимеразами. По названию затравки – праймер эти полимеразы получили название праймаз). Все ДНК-полимеразыформируют дочерние цепи в одном направлении 5’ à 3’, перемещаясь по матричной ДНК в направлении 3’ à 5’. На ДНК-полимеразывозложена ответственность за точность репликации, поэтому многие из них обладают экзонуклеазной корректирующей активностью, действующей в направлении 3’à 5’. Субстратами ДНК полимераз являются дезоксирибонуклеозидтрифосфаты.Инициация репликации ДНК начинается в специальных участках со специфической нуклеотидной последовательностью (Эти участки получили название ori от origin - начало) и продолжается в двух направлениях до тех пор, пока репликативные вилки не встречаются на противоположной стороне кольцевой хромосомы бактерии. После завершения процесса две связанные кольцевые дочерние молекулы разделяются при участии специальной ферментной системы.. Скорость репликации (перемещения вилки) у бактерий составляет приблизительно 50,000-100000 пар нуклеотидов в минуту при 37 0 C. Родительская двойная спираль ДНК при такой скорости перемещения должна раскручиваться со скоростью 5,000-10000 об\мин (один виток молекулы образуется 10 парами азотистых оснований). Такой простой расчет указывает на определенные чисто механические проблемы, возникающие в процессе репликации. Основные участники процесса репликации и их основные функции у прокариот перечисляются в следующей таблице.
Последовательность событий в механизме репликации у Е Coli следует строго определенному сценарию. oric - инициирующая последовательность E.Coli длиной в 245 пар азотистых оснований имеет участок с четырьмя повторами последовательности из 9- пар оснований и трех повторов из 13 пар оснований. Последние характеризуются высоким содержанием АТ пар и таким образом денатурируются относительно легко. К этому участку присоединяется белок - продукт гена DnaA.и специфические основные белки (HU и IHF) которые облегчают изгиб ДНК, образование которого - важный шаг в инициации репликации:
атем в дело вступает ДНК-полимераза III. Как оказалось, выше упомянутая ДНК полимераза открытая А.Корнбергом и теперь называемая ДНК-полимеразаI (Pol I) - не основной фермент репликации, хотя и играет существенную роль в процессе репликации (см. позже). У E. coli выделены две дополнительных ДНК-полимеразы, полимераза II (Pol II) и полимераза III (Pol III). Точная функция полимеразы II в клетке пока не известна. Основным ферментом, катализирующим репликацию, является полимераза III. Помимо полимеразной активности все бактериальные ДНК-полимеразыобладают 3 '-5 ' экзонуклеазной активностью. Способность катализировать гидролиз фосфодиэфирных связей и удалять нуклеотиды с 3’ концов, действуя в противоположном от своей полимеразной активности направлении, позволяет полимеразам проверять и устранять ошибки своей деятельности.
Если к растущей цепи добавляется неправильный нуклеотид, он немедленно удаляется до завершения синтеза. У полимеразы I и III обнаружена и 5 '-3 ' экзонуклеазная активность, что позволяет им участвовать в процессах репарации ДНК. Эта же экзонуклеазная активность полимеразы I используется также для удаления РНК- затравок, которые инициируют синтез цепей ДНК (см. ниже). Работе ДНК- полимеразы предшествует первичный отбор комплементарного матрице дНТФ, который проходит в общем одинаково для разных ДНК-полимераз Этот отбор начинается с образования канонических Уотсон-Криковских пар оснований. Их образование энергетически более выгодно, поскольку при этом образуется оптимальное по сравнению с другими возможными вариантами число водородных связей. Кроме того, правильно образованные пары оказывают влияние на конформационное изменение ДНК-полимеразы, направленное на ускорение реакции элонгации и последующего перемещения фермента. При образовании неправильной пары резко ухудшается кинетика процесса. Скорость перемещения фермента замедляется. При кинетической задержке элонгации цепи, возникающей в результате включения некомплементарного 3'-нуклеотидного остатка и возникновения ошибочной пары начинает проявляться активность другого центра ДНК полимеразы, расположенного вблизи центра полимеризации и, по-видимому, частично совмещающегося с ним. Этот активный центр, катализирующий 3'->5'-экзонуклеазный гидролиз удаляет некомплементарный 3'-концевой нуклеотидный остаток. Ошибочные пары возникают с частотой порядка 1: 104 - 1: 106. Дело в том, что ДНК-полимеразе на каждом шагу элонгации дДНК приходится выбирать один из четырех различающихся по структуре дНТФ в условиях, когда матричный нуклеотидный остаток в образующейся паре также вариабелен из-за соседних нуклеотидных остатков, влияющих на конформацию односпирального участка матрицы в активном центре. Редактирующая активность фермента понижает частоту ошибок в 10-100 раз. ДНК-полимераза копирует антипараллельные цепи ДНК перемещаясь в 5 'à 3 ' направлении Как можно объяснить, что ДНК-полимераза копирует антипараллельные цепи матрицы ДНК в одном 5 'à 3 ' направлении одновременно? Предложена модель, согласно которой ДНК-полимераза формирует димер, связанный с другими необходимыми в репликативной вилке белками (реплисома). Одна из цепей матричной ДНК, названная отстающей цепью, временно образует петли вокруг реплисомы так, что димер ДНК-полимеразы получает возможность перемещаться по обеим цепям в одном 3' à 5 ' направлении одновременно на короткое расстояние. Мономер ДНК-полимеразы, копирующий эту цепь обладает низкой процессивностью и через небольшой промежуток времени, синтезировав короткий отрезок ДНК, покидает матричную отстающую цепь. Эти короткие (до 1000 пар нуклеотидов) отрезки ДНК, образующиеся на отстающей цепи, получили название фрагментов Оказаки по имени Ф. Оказаки, впервые (1968) указавшего на прерывистый характер синтеза ДНК. Новое присоединение мономера ДНК-полимеразык матрице происходит после предварительного синтеза нового праймера. Вторая половина димера ДНК-полимеразыреплисомы, обладающая более высокой процессивностью, в чем ей помогают специфические белки, копирует дочернюю цепь ДНК без перерывов. Матричную цепь, которая копируется без перерывов, назвали ведущей цепью. Так как репликативная вилка довольно быстро (1000 нулеотидов в сек) продвигается по матрице вновь синтезируемые дочерние цепи и родительские цепи матрицы сразу формируют двойные спирали ДНК. Предполагается, что только маленький отрезок матричной двойной спирали находится в одноцепочечном состоянии в данный отрезок времени. Праймеры ведущей и отстающей цепей (10-12 пар нуклеотидов) удаляются ДНК полимеразой I, обладающей репарирующей функцией с одновременной заменой рибонуклеотидов дезоксирибонуклеотидами. Промежутки, которые возникают между 3 '-ОН и 5 '- фосфатом, воссоединяются лигазами ДНК, завершая тем самым процесс репликации.
Ретровирусы внесли изменения в центральную догму молекулярной биологии. Особой формой репликации у прокариот является репликация с использованием РНК в качестве матрицы для синтеза ДНК. Такая форма репликация обнаружена у ретровирусов (например, у HIV ретровируса, вызывающего СПИД) и катализируется ферментом РНК зависимой ДНК полимеразой или о братной транскриптазой. Подобно другим РНК зависимым полимеразам, обратная транскриптаза - обладает высокой склонностью к ошибкам, посколько не обладает коррегирующей активностью. Схема репликации РНК ретровирусов следующая: 1. Вирусная РНК в клетке хозяина образует пары оснований со специфической молекулой тРНК, образуя праймер для репликации ДНК. 2. Обратная транскриптаза синтезирует ДНК с образованием смешаной ДНК-РНК молекулы. 3. Рибонуклеаза H частично разрушает цепь РНК двойной РНК-ДНК спирали, удаляя 5 ' концевой отдел. 4. 3 ' конец вирусной РНК образует пары с освободившейся цепью ДНК, формируя кольцевую структуру. 5. Обратная транскриптаза создает копию цепи ДНК генома, используя 3 ' конец цепи ДНК в качестве праймера. 6. Оставшаяся цепь РНК удаляется, и вновь синтезированная цепь ДНК образует комплементраную двойную спираль с первой цепью ДНК. 7. Образовавшаяся полная молекула ДНК внедряется в хромосому клетки- хозяина
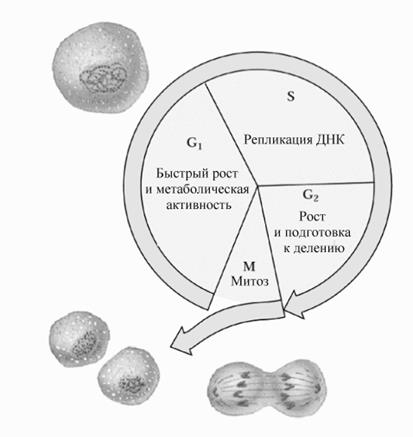
Препарат 3 '-азидо-2 ' 3 '-дидезокситимидин (АZT), после превращения в соответствующий 5 ' трифосфат в клетках, является ингибитором фермента обратной транскриптазы вируса HIV. Другие аналоги нуклеозидов, типа 2 ' 3 '-дидезоксицитидин (ддЦ), 2 ' 3 '-дидезоксиинозин и 2 ' 3 '-дидегидро-3 '-дезокситимидин (d4T) после фосфорилирования в соответствующие трифосфаты блокируют удлинение цепи ДНК после включения в ДНК. . Биосинтез ДНК у эукариот связан с циклом деления клетки.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 427; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.205.149 (0.013 с.) |