Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Системи схрещувань організмів та їхні генетичні наслідкиСодержание книги
Поиск на нашем сайте
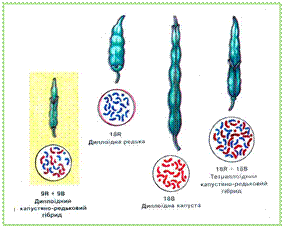
Гібридизація можлива між особинами одного (внутрішньовидова) та різних видів з одного чи двох родів (міжвидова, або віддалена) організмів. Внутрішньовидове схрещування буває спорідненим і неспорідненим. Споріднене схрещування, або інбридинг (від англ. ін, - в, усередині та бридінг - розведення) — це схрещування організмів, що мають безпосередніх спільних предків. Залежно від ступеня спорідненості інбридинг може бути більш або менш тісним. Найтісніші форми спорідненого схрещування спостерігають серед самозапильних рослин і самозаплідних гермафродитних тварин. У організмів з перехресним заплідненням тісний інбридинг відбувається при схрещуванні братів і сестер, батьків та їхніх нащадків тощо. Унаслідок спорідненого схрещування з кожним наступним поколінням підвищується гомозиготність гібридів. Це пояснюється тим, що генетична близькість батьків збільшує ймовірність поєднання у потомках одних і тих самих алелів різноманітних генів пропорційно до ступеня спорідненості. У самозапильних рослин уже у 10-му поколінні спостерігається майже повна гомозиготність (до 99,9%), а при схрещуванні братів із сестрами чи батьків з нащадками такий результат може бути досягнутий після 20-го покоління Проте досягнути 100%-ї гомозиготності за всіма генами не вдається, бо вона порушується внаслідок мутацій, що постійно виникають. Біологічним наслідком спорідненого схрещування є ослаблення або навіть виродження нащадків, оскільки у таких гомозиготних організмів збільшується ймовірність фенотипного прояву летальних чи сублетальних рецесивних алелей. Таким чином, тісний інбридинг часто призводить до появи організмів із різноманітними спадковими аномаліями. Наслідки спорідненого схрещування були відомі людині ще в сиву давнину. Наприклад, близько 20% людей-альбіносів є нащадками від споріднених шлюбів. У людини є декілька рецесивних летальних алелей, здатних у гомозиготному стані спричинити смерть. Тому шлюби між близькими родичами вважалися небажаними або взагалі заборонялися релігією чи законами. У селекції споріднене схрещування застосовують для отримання чистих ліній. Інбридинг дає можливість дістати цінні ознаки в гомозиготному стані і закріпити їх серед нащадків. Неспоріднене схрещування, або аутбридинг (від англ. аут — поза) — гібридизація організмів, які не мають тісних родинних зв'язків, тобто представників різних ліній, сортів чи порід одного виду. Неспорідненими вважають особин, у яких немає спільних предків принаймні протягом попередніх шести поколінь. Неспоріднене схрещування застосовують для поєднання у потомстві цінних властивостей, притаманних різним лініям, породам чи сортам. За своїми генетичними наслідками воно прямо протилежне інбридингу. При неспорідненому схрещуванні з кожним наступним поколінням підвищується гетерозиготність нащадків. Це пояснюється тим, що зі зменшенням ступеня спорідненості організмів зростає ймовірність наявності в них різних алелів певних генів. У разі неспорідненого схрещування часто спостерігається явище гетерозису, або «гібридної сили». Гетерозис (від грец. гетероіозіс - зміна, перевтілення) - це явище, за якого перше покоління гібридів, одержаних у результаті неспорідненого схрещування, має підвищені життєздатність і продуктивність порівняно з вихідними батьківськими формами. Це пояснюється тим, що у гетерозисних форм сублетальні та летальні рецесивні алелі переходять у гетерозиготний стан, завдяки чому їхня шкідлива дія не проявляється фенотипно. Крім того, в генотипі гібридних нащадків можуть поєднуватися сприятливі домінантні алелі обох батьків. Унаслідок цього може спостерігатися явище взаємодії неалельних домінантних генів. За даними біохімічних досліджень у гетерозисних форм часто спостерігається ширший набір ферментів та їхня підвищена активність порівняно з батьківськими організмами. Гетерозис повною мірою проявляється в першому поколінні гібридів, однак у наступних, унаслідок розщеплення ознак та переходу частини генів у гомозиготний стан, його ефект слабшає і до восьмого покоління сходить нанівець. У рослин гетерозис можна закріпити вегетативним розмноженням, подвоєнням числа хромосом або партеногенезом. Гетерозис може більше позначитись на одних варіантах ознак гібридної особини, не зачіпаючи інших. Явище гетерозису широко застосовують у сільському господарстві, оскільки воно значно підвищує продуктивність (наприклад,у кукурудзи - до 20-25%), в овочевих культур (цибулі, помідорів, огірків, баклажанів, буряків тощо). У тваринництві схрещування між різними породами прискорює рістістатеве дозрівання, поліпшує якість м'яса, молока тощо. Так, у результаті схрещування несучих порід курей між собою (наприклад, леггорнів з австралорнами) продуктивність гібридів зростає на 20-25 яєць у рік. У бройлерів (гібридних курчат м'ясних порід) гетерозис прискорює ріст і поліпшує якість м'яса. Віддалена гібридизація - схрещування особин, які належать до різних видів і навіть родів з метою поєднання у гібридів цінних спадкових ознак представників різних видів. За допомогою віддаленої гібридизації створено гібриди пшениці та пирію, що відзначаються високою продуктивністю (до 300-450 ц/га зеленої маси)тастійкістю до полягання; пшениці з житом (тритікале); китайської цукрової тростини з дикими видами, що сприяло підвищеннюцукристості. Відомі міжвидові гібриди і серед плодових культур (малинита ожини, сливи й терену, горобини та сибірського глоду тощо). У тваринництві також виведено значну кількість міжвидових гібридів. Наприклад, гібрид кобили та осла - мул, який відрізняється значною витривалістю, фізичною силою та довшим терміном життя порівняно з батьківськими формами. Подібні властивості виявляє гібрид одногорбого та двогорбого верблюдів. Унаслідок міжвидового схрещування виведено нову породу овець - архаромериносів (гібрид тонкорунних овець - мериносів і дикого гірського барана - архара). Схрещуванням свійської тварини високогірських районів Центральної Азії - Яка — з великою рогатою худобою виведено нові породи (сарлики та хайнаки). Вони мають більшу продуктивність і вищі якості м'яса порівняно з яком та кращу пристосованість до суворих умов високогір'я порівняно з великою рогатою худобою. У рибництві відомий гібрид білуги та стерляді — бістер, який швидко росте та має смачне м'ясо. Селекціонери часто стикаються з проблемою безпліддя міжвидових гібридів, гамети яких звичайно не дозрівають. Навіть за умов однакової кількості хромосом у каріотипах батьківських форм хромосоми різних видів різняться за структурою і тому нездатні до кон'югації. Особливо ускладнюється хід мейозу за умов різного числа хромосом у каріотипах батьків. Так, мули нездатні до розмноження; у гібрида Яка та великої рогатої худоби самці безплідні, а самки – плодючі. Уперше методику подолання стерильності міжвидових гібридів у рослин розробив у 1924 році Г.Д. Карпеченко на прикладі гібрида капусти та редьки, який за своїм фенотипом займав проміжне положення між відповідними фенотипами батьків (мал.39.3.).
Мал. 39.3. Подолання безпліддя міжвидового капустяно-редькового гібрида на основі поліплоїдії Хоча капуста і редька належать до різних родів родини капустяних (хрестоцвітих), вони мають однакову кількість хромосом (2n = 18). Утім, створений Г.Д.Карпеченком міжвидовий гібрид виявився безплідним, оскільки під час мейозу «капустяні» та «редькові» хромосоми між собою не кон'югували. Тоді вчений подвоїв хромосомний набір гібрида (4 n = 36), тобто в ядрі його клітин тепер було по два повні набори хромосом кожного з батьків (по 2 «капустяні» та 2 «редькові» гомологічні хромосоми кожної пари). Унаслідок цього процес мейозу у такої поліплоїдної форми перебігав нормально: «капустяні» хромосоми кон'югували з «капустяними», а «редькові» — з «редьковими» і в кожну з гамет завжди потрапляло по одному гаплоїдному набору хромосом редьки та капусти.
|
|||||||
|
|
Последнее изменение этой страницы: 2016-07-16; просмотров: 1530; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.17.181.181 (0.01 с.) |