Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механизм мышечного сокращенияСодержание книги
Поиск на нашем сайте
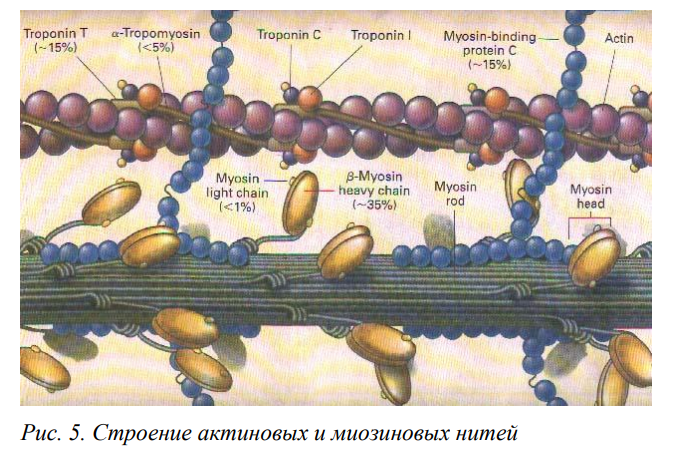
В 1954 г. Хаксли показал, что при мышечном сокращении миозиновые и актиновые нити не изменяют своей длины. Изменение саркомера происходит за счет взаимного перекрывания актина и миозина, т.е. скольжения нитей. В присутствии АТФ, но при низкой концентрации Са2+, волокно находится в расслабленном состоянии, т.е. головки миозина не образуют связи с актином. Соединение актина и миозина препятствуют тропонин, который «закрывает» участки связывания на актине. При возбуждении мышечного волокна ионы кальция выходят из СПР и его концентрация вблизи миофибрилл возрастает. В момент увеличения концентрации Са2+ вблизи головок ферментативная активность головок многократно увеличивается и в той же мере возрастает гидролиз АТФ. В скелетных мышцах тяжи тропомиозина закрывают активные участки актинового филамента, препятствуя взаимодействию миозиновых головок с мономерами актина, тем самым предотвращая сокращение. Повышение концентрации Са2+ сопровождается его связыванием с тропонином. При этом молекулы комплекса тропонин-тропомиозин изменяют свое расположение таким образом, что обнажают активные участки на актиновом филаменте. За счет освобождающейся энергии происходит связывание головок с нитью актина, изгибание «ножки» и вследствие этого актиновые филаменты продольно перемещаются относительно центрального миозинового стержня. Уменьшение же концентрации Са2+ до прежнего минимального уровня вызывает размыкание поперечных мостиков и возвращение мышечного волокна (всей мышцы) в исходное расслабленное состояние.
Описанное взаимодействие глобулярной головки миозина и активным участком молекулы актина называют циклом поперечного мостика. Один цикл поперечного мостика происходит в следующей последовательности: -активация миозиновой головки; -обнажение активного участка молекулы актина в присутствии кальция; -самопроизвольное формирование поперечного мостика; -поворот глобулярной головки, сопровождающийся продвижением актиновой нити и укорочением саркомера; Цикл после завершения может повториться или остановиться. Если бы все поперечные мостики в отдельной мышце прошли одновременно только один цикл, мышца укоротилась бы всего на 1% от ее длины в состоянии покоя. Однако многие мышцы способны укорачиваться до 60% длины покоя. Это показывает, что цикл поперечного мостика происходит в них многократно. При этом каждый раз миозин захватывает и тянет новый участок актина. Именно такая многократность повторения цикла замыкания-размыкания мостика вызывает более выраженное укорочение и генерирует большую силу. В результате циклов поперечных мостиков саркомер укорачивается, и концы мышцы подтягиваются к ее центру. Прочная соединительнотканная фасциальная оболочка, прикрепляющая мышцу посредством сухожилия к костям, передает вызываемую сокращением силу костям. Линейно направленная мышечная сила тянет кость и вызывает ее вращение в плоскости, перпендикулярной оси сустава. Поскольку мышца обычно проходит, по крайней мере, через один сустав, более свободный сегмент костного скелета будет перемещаться, вызывая вращательное движение в суставе.
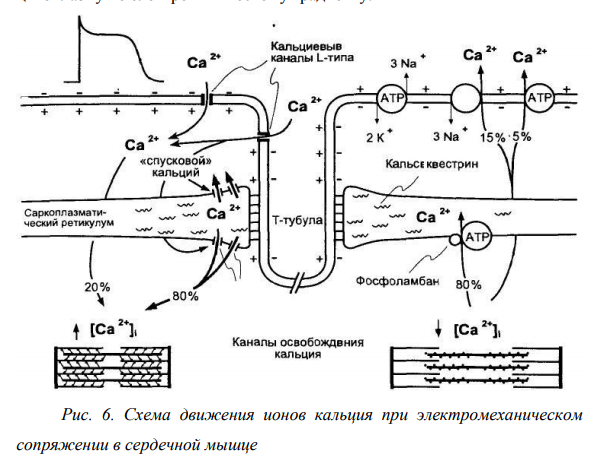
Трансформация возбуждения в сокращение, по существу представляющая передачу сигнала о начале сокращения от возбужденной сарколеммы, называется электромеханическим сопряжением. Электромеханическое сопряжение происходит посредством распространения возбуждения (потенциала действия) по системе поперечных трубочек внутрь волокна. При этом информация о возбуждении (фактически о перезарядке мембраны) очень быстро достигает продольной системы СПР, в конечном итоге вызывая открытие в мембране цистерн Са2+-каналов и залповый выброс катиона кальция, быстро диффундирующего к миофибриллам. Последующее связывание головок миозина с актином вызывает напряжение волокна или генерацию силы. Высвобождение Са2+ из СПР поперечно-полосатых мышц во время сопряжения возбуждения и сокращения происходит благодаря взаимодействию Т-трубочек и концевых цистерн СПР. Предполагают, что развивающаяся при передаче потенциала действия по Т-трубочке деполяризация вызывает конформационные изменения белков ножки концевой цистерны. Эти конформационные изменения приводят к открытию Са2+ -каналов в СПР и обеспечивают выход ионов Са2+ в цитоплазму по электрохимическому градиенту.
Рис. 6. Схема движения ионов кальция при электромеханическом сопряжении в сердечной мышце
Сигналом к остановке сокращения скелетной мышцы служит прекращение нервного импульса в двигательной концевой пластинке нервно-мышечного соединения. Прекращение импульса означает устранение сигнала к высвобождению кальция. Циклическое образование поперечных мостиков продолжается до тех пор, пока в волокне имеются в достаточном количестве свободных катионов кальция и АТФ. Все, что снижает концентрацию Са2+ и АТФ, будет ослаблять мышечное сокращение, что происходит при утомлении мышцы. Таковыми факторами, например, является низкая концентрация калия, яды и токсины, нарушающие выработку АТФ в окислительном фосфорилировании.
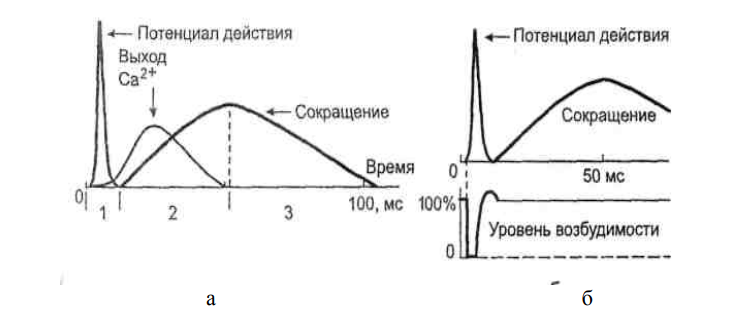
Полное отсутствие АТФ вызывает устойчивое сокращение, называемое трупным окоченением. Такое состояния устойчивого сокращения обусловлено двумя обстоятельствами. Во-первых, в отсутствие АТФ невозможна откачка Са2+ и саркоплазмы в везикулы СПР. Во-вторых, без АТФ не может произойти повторная активация миозиновой головки, требующаяся для разрыва поперечного мостика между актином и миозином. В итоге поперечные мостики остаются замкнутыми, а мышца в сокращенном состоянии. Выделяют два вида мышечных сокращений: одиночное и тетаниче- ское. Одиночное мышечное сокращение проявляется в результате развития одиночной волны возбуждения в мышечных волокнах. Этого можно достичь при воздействии на мышцу очень коротким (около 1 мс) стимулом. В развитии одиночного мышечного сокращения выделяют латентный период, фазу укорочения и фазу расслабления. Сокращение мышцы начинает проявляться через 10 мс от начала воздействия раздражителя. Этот временной интервал называют латентным периодом (рисунок 1). Затем последует развитие укорочения (длительность около 50 мс) и расслабления (50-60 мс). Считается, что на весь цикл одиночного мышечного сокращения затрачивается в среднем 0,1 с. Но следует иметь в виду, что длительность одиночного сокращения у разных мышц может сильно варьировать. Она также зависит от функционального состояния мышцы. Скорость сокращения и особенно расслабления замедляется при развитии утомления мышцы. К быстрым мышцам, имеющим короткий период одиночного сокращения, относятся мышцы языка и смыкающие веко.
Рисунок 1 – Временные соотношения разных проявлений возбуждения волокна скелетной мышцы: а – соотношение потенциала действия, выхода Ca2+ в саркоплазму и сокращения: 1 – латентный период; 2— укорочение; 3 - расслабление; б – соотношение потенциала действия, со- кращения и уровня возбудимости. Под влиянием одиночного раздражителя вначале возникает потенциал действия, и лишь затем начинает развиваться период укорочения. Оно продолжается и после окончания реполяризации. Восстановление исходной поляризации сарколеммы свидетельствует и о восстановлении возбудимости. Следовательно, на фоне развивающегося сокращения в мышечных волокнах можно вызвать новые волны возбуждения, сократительный эффект от которых будет суммироваться. Тетаническим сокращением или тетанусом называют сокращение мышцы, появляющееся в результате возникновения в моторных единицах многочисленных волн возбуждения, сократительный эффект от которых суммируется по амплитуде и времени. Различают зубчатый и гладкий тетанус. Для получения зубчатого те- тануса надо стимулировать мышцу с такой частотой, чтобы каждое после- дующее воздействие наносилось после фазы укорочения, но до момента окончания расслабления. Гладкий тетанус получается при более частых раздражениях, когда последующие воздействия наносятся во время разви- тия укорочения мышцы. Например, если фаза укорочения у мышцы составляет 50 мс, а фаза расслабления 60 мс, то для получения зубчатого те- тануса необходимо раздражать эту мышцу с частотой 9-19 Гц, для полу- чения гладкого – с частотой не менее 20 Гц.
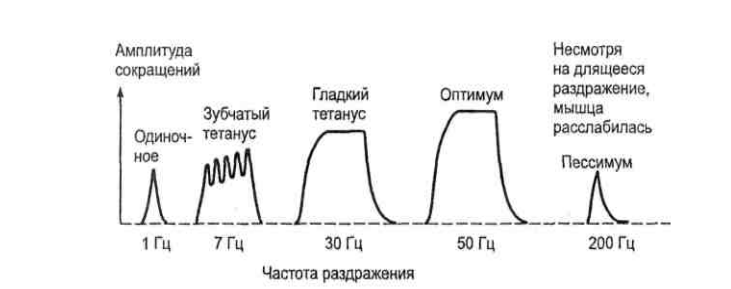
Для демонстрации различных видов тетануса обычно используют ре- гистрацию сокращений изолированной икроножной мышцы лягушки на кимографе. Пример такой кимограммы представлен на рисунке 2. Ам- плитуда одиночного сокращения минимальна, увеличивается при зубча- том тетанусе и становится максимальной – при гладком. Одной из причин такого возрастания амплитуды является то, что при возникновении час- тых волн возбуждения в саркоплазме мышечных волокон накапливается Ca2+, стимулирующий взаимодействие сократительных белков.
Рисунок 2 – Зависимость амплитуды сокращения от частоты раз- дражения (сила и длительность стимулов неизменны) При постепенном увеличении частоты раздражения нарастание силы и амплитуды сокращения мышцы идет лишь до некоторого предела – оп- тимума ответной реакции. Частоту раздражения, вызывающую наи- больший ответ мышцы, называют оптимальной. Дальнейшее увеличение частоты раздражения сопровождается уменьшением амплитуды и силы сокращения. Это явление называют пессимумом ответной реакции, а частоты раздражения, превышающие оптимальную величину, – пессимальными. Явления оптимума и пессимума были открыты Н.Е. Введенским. При оценке функциональной активности мышц говорят об их тонусе и физических сокращениях. Тонусом мышцы называют состояние длитель- ного непрерывного напряжения. При этом видимое укорочение мышцы может отсутствовать из-за того, что возбуждение возникает не во всех, а лишь в некоторых моторных единицах мышцы и они возбуждаются не синхронно. Физическим сокращением мышцы называют кратковременное укорочение мышцы, сменяющееся ее расслаблением.
На одиночный стимул мышца отвечает одиночным мышечным сокращением. Раздражение, наносимое на мышцы характеризуется следующими параметрами: 1) длительностью, 2) интенсивностью, 3) частотой. Длительность единичного мышечного сокращения составляет примерно 0,1 с. Если на мышцу наносить повторные раздражения постоянной силы и длительностью с частотой, несколько ниже той, при которой возникает тетанус, то можно наблюдать феномен лестницы.
Электрический ответ мышцы на раздражение характеризуется периодом рефрактерности, когда мышца не отвечает на раздражение, в механическом же сокращении скелетной мышцы такого периода нет. Поэтому если на мышцу наносить повторное раздражение в тот момент, когда она еще не полностью расслабилась после предшествующего сокращения, можно наблюдать усиление сокращения, или суммацию. Длительность напряжения мышцы приводит к еѐ утомлению и неспособности совершать дальнейшую работу. Поступление нервных импульсов к мышце может быть нормальным, однако механическое сокращение мышцы в ответ на эту импульсацию угнетено вследствие истощения основного источника энергии – АТФ.
|
||||||||
|
|
Последнее изменение этой страницы: 2022-09-03; просмотров: 57; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.117.141.116 (0.008 с.) |