Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Кальций является вторичным мессенджером для многих сигнальных путей⇐ ПредыдущаяСтр 20 из 20
Существует множество вариантов сигнализации с участием ионов кальция. Во многих клетках, отвечающих на внеклеточные сигналы, Са2+ служит вторичным мессенджером, приводящим в действие внутриклеточные ответы, такие как экзоцитоз в нейронах и клетках эндокринной системы, сокращение мышц и перестройку цитоскелета в процессе амебоидного движения. В норме концентрация Са2+ в цитозоле сохраняется очень низкой (<10 7 М) благодаря действию Са2+-насосов в эндоплазматическом ретикулуме ЭР, митохондриях и плазматической мембране. Гормональные сигналы, нервные импульсы или другие стимулы вызывают либо вход Са2+ в клетку через специфические Са2+-каналы в плазматической мембране, либо высвобождение кальция из ЭР или митохондрий, в каждом случае происходит увеличение содержания Са2+ в цитозоле и запуск клеточного ответа. Изменения концентрации Са2+ воспринимаются Са2+-связывающими белками, регулирующими множество Са2+-зависимых ферментов. Кальмодулин (СаМ; Мг = 17000) — кислый белок с четырьмя высокоаффинными Са2+-связывающими центрами. Когда содержание Са2+ в клетке повышается до -106 М (1 мкМ), присоединение Са2+ к кальмодулину вызывает в этом белке конформационное изменение.
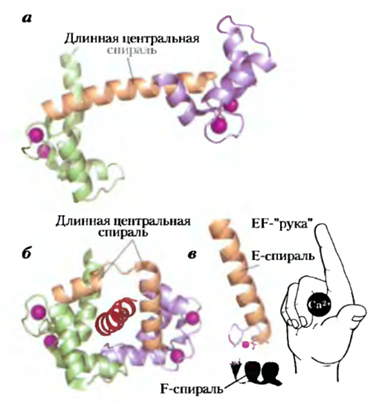
Рис. Кальмодулин — белковый посредник для многих ферментативных реакций, стимулируемых ионами Са2+. Кальмодулин содержит четыре высокоаффинных Са2+-связывающих центра (Кй ~ 0,1 — 1 мкМ). а) Ленточная модель кристаллической структуры каль- модулина (PDB ID 1CLL). Четыре Са2+-связывающих центра заняты ионами кальция (фиолетовый). Слева — N-концевой домен; справа — С-концевой домен. б) Кальмодулин, связанный со спиральным доменом (красный цвет) одного из многих белков, которые он регулирует, — кальмодулинзависимой протеинкиназой II (PDB ID 1CDI). Заметим, что длинная центральная а- спираль, видимая на (а), изгибается назад при связывании со спиральным доменом субстрата. Центральная спираль, разумеется, в растворе более подвижна, чем в кристалле, в) Каждый из четырех Саг+-связывающих центров находится во фрагменте спираль-петля-спираль, называемом «EF-рукой», который присутствует также во многих других Са2+-связывающих белках. Кальмодулин связывается с множеством белков и при присоединении к нему Са2+ модулирует их активность. Кальмодулин — член семейства Са2+-связывающих белков, туда же относится тропонин, инициирующий сокращение скелетных мышц в ответ на увеличение концентрации ионов кальция. Для этого семейства характерна Са2+-связывающая структура — «EF-рука». Кальмодулин является также интегральной субъединицей семейства ферментов, Са2+/ кальмодулинзависимых протеинкиназ (СаМ-киназы I—IV). Когда в ответ на какой-либо стимул внутриклеточная концентрация Са2+ увеличивается, кальмодулин связывает Са2+, претерпевает конформационное изменение и активирует СаМ-киназу. Эта киназа затем фосфорилирует ряд ферментов-мишеней, регулируя их активность.
Кальмодулин служит также регуляторной субъединицей мышечной киназы фосфорилазы Б, которая активируется ионами Са2+. Таким образом, Са2+ вызывает АТР- зависимые сокращения мышц, активируя в то же время и распад гликогена, обеспечивающий «топливом» синтез АТР. Известно, что многие другие ферменты тоже регулируются ионами Са2+ при помощи кальмодулина. Активность Са2+ как вторичного мессенджера, подобно активности сАМР, можно ограничить на какой-либо территории; после того как его высвобождение вызвало локальный ответ, Са2+ обычно удаляется, прежде чем диффундирует в отдаленные части клетки. Очень часто концентрация Са2+ не просто увеличивается и потом уменьшается, а, скорее, колеблется с периодом в несколько секунд, даже когда концентрация гормона снаружи клетки остается постоянной. Механизм, лежащий в основе колебания концентрации ионов кальция, по-видимому, связан с регуляцией по типу обратной связи либо фосфолипазы, которая генерирует IP3, либо ионного канала, который отвечает за высвобождение Са2+ из ЭР, либо и того, и другого. Каким бы ни был механизм, эффект заключается в том, что один вид сигнала (например, концентрация гормона) превращается в другой (частота и амплитуда колебаний внутриклеточного содержания Са2+). Другим возможным изменением являются локальные «пятна», «облака» или «волны» ионов кальция — временное увеличение концентрации ионов Са2+, ограниченные определенным отделом клетки. Сигнал Са2+ ослабевает по мере того, как ионы кальция диффундируют от источника (кальциевого канала), удерживаются в ЭР или выкачиваются из клетки. Сигнальные системы с участием Са2+ и с AMP связаны между собой. В некоторых тканях ионы кальция стимулируют как аденилатциклазу, производящую с AMP, так и фосфодиэстеразу, разрушающую с AM Р. Следовательно, временные и локальные изменения концентрации ионов кальция могут приводить к временным и локальным изменениям концентрации сАМР. Мы уже упоминали, что сАМР-зависимый фермент РКА часто входит в состав строго локализованного надмолекулярного комплекса, образующегося вокруг каркасных белков, таких как АКАР. Внутриклеточная локализация ферментов-мишеней в сочетании с временным и пространственным градиентами концентрации Са2* и с AMP позволяет клетке отвечать на один или несколько сигналов соответствующими метаболическими изменениями, локализованными во времени и в пространстве.
Мембранный фосфолипид Р I Р3 работает в одной из ветвей передачи сигнала инсулина!!! PIP 2 Фосфатидилинозит и его фосфорилированные производные участвуют в нескольких уровнях регуляции клеточной структуры и метаболизма. Фосфатидилинозит-4,5-бисфосфат на цитоплазматической (внутренней) поверхности плазматической мембраны является запасной формой молекул-посредников, высвобождающихся внутрь клетки в ответ на внеклеточные сигналы, действующие на специфические рецепторы, расположенные на внешней поверхности плазматической мембраны. Внеклеточные сигналы, такие как гормон вазопрессин, активируют фосфолипазу С в мембране. Она гидролизует фосфатидилинозит-4,5-бисфосфат, высвобождая два продукта, которые действуют как внутриклеточные мессенджеры: инозит-1,4,5- трисфосфат (IP3). который растворим в воде, и диацилглицерин, который остается в связанном с плазматической мембраной виде. Молекула IР3 инициирует выход Са2* из эндоплазматического ретикулума, а комбинация диацилгицери- на и увеличенной концентрации Са2~ в цитозоле активирует фермент протеинкиназу С. Этот фермент фосфорилирует специфические белки, что вызывает клеточный ответ на внеклеточный сигнал. Такой сигнальный механизм более подробно описан в гл. 12 (см. рис. 12-10). Инозитные фосфолипиды также служат в качестве «зародышей» для образования некоторых надмолекулярных комплексов, участвующих в передаче сигналов или в экзоцитозе. Специальные сигнальные белки специфически связываются с фосфатидилинозит-3,4,5-трисфосфатом в плазматической мембране, инициируя образование мультиферментных комплексов на обращенной в цитозоль поверхности мембраны. Образование фосфатидилинозит-3,4,5-трисфосфата в ответ на внеклеточные сигналы удерживает белки вместе в сигнальных комплексах на поверхности плазматической мембраны (см. рис. 12-16). Мембранные сфинголипиды могут также служить источником внутриклеточных мессенджеров. И церамид, и сфингомиелин — мощные регуляторы протеинкиназ; известно, что церамид или его производные участвуют в регуляции клеточного деления, диффе- ренцировки, миграции и программируемой клеточной гибели (апоптоза).
Если вспомнить биохимию😊
Лекция 9
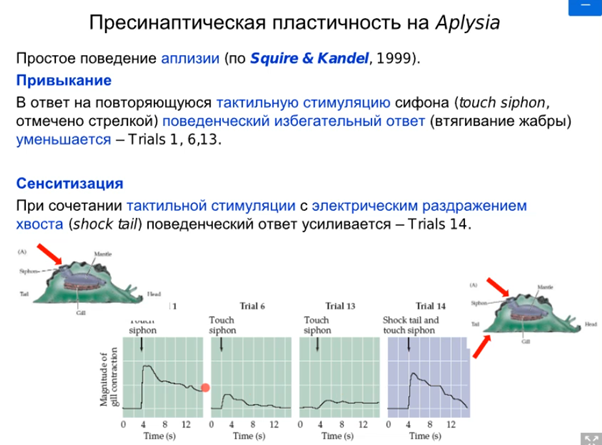
Напоминание от Латанова про несчастную аплизию и что есть не ассоциативные формы научения
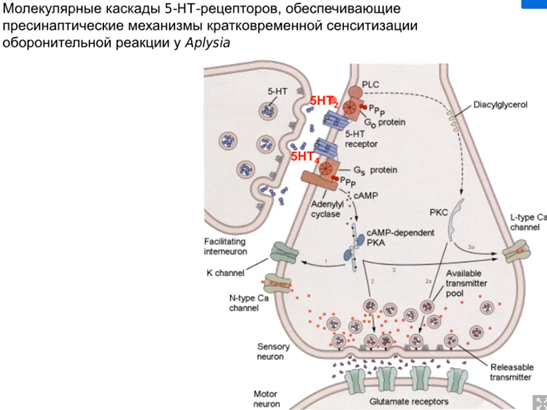
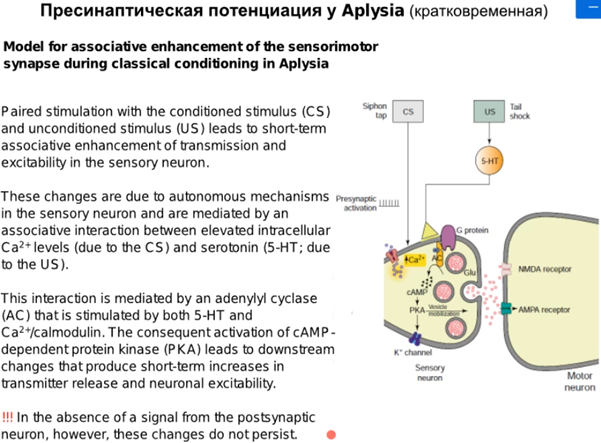
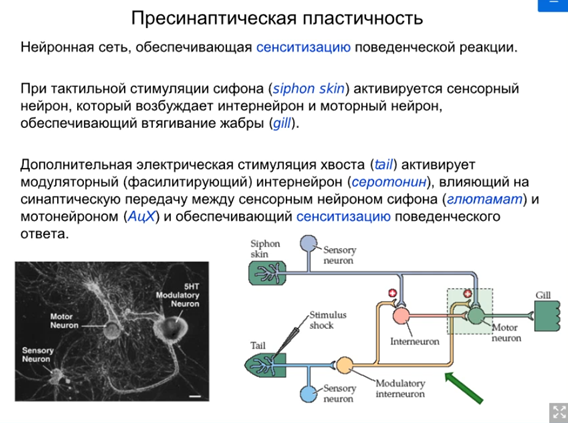
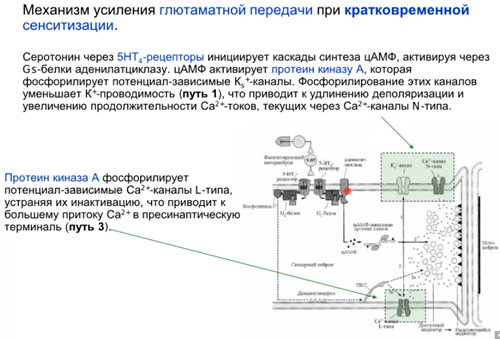
Нас интересует аксоаксональный синапс от модуляторного нейрона от хвоста Ниже картинка того, что происходит в аксонной терминали сенсорного нейрона сифона(1). Зеленой стрелкой справа кратко показаны слева направо хвост ->сенсор нейрон(2)-> фасилитирующий серотониновый(который модуляторный) интернейрон, который из своей терминали выделяет серотонин и он попадает на терминаль аксона сенсорного нейрона, где есть два вида 5-HT рецепторов (2 и 4 тип). Ниже каскады
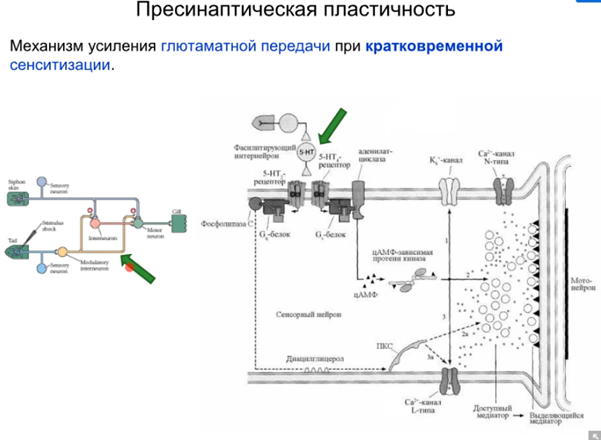
Просто есть путь 1(открыт первый це классика, см картинку про всякую химозу) и путь 3. Первый путь фосфорилирует в результате калиевые каналы, зависящие от серотонина (Кs каналы), что уменьшает их проводимость (калиевые токи всегда тормозные и эти каналы всегда открыты), что приводит к увеличению времени кальциевого тока и большему количеству везикул в терминали (помним что в везикулах есть синаптотагмин - кальциевый сенсор, который запускает экзоцитоз). Комментарий от Латанова (любая проводимость на мембране нейрона уменьшает сопротивление U= RI, то есть нужны большие токи чтобы достигнуть уровня срабатывания потенциал зависимых каналов, потом латанов сказал все тоже самое и сказал что нужны почему то МЕНЬШИЕ токи но я хз, здравый смысл говорит, что в начале он правильно сказал). Осознав самостоятельно отдельно от Латанова и почитав википедию, я пришла к выводу, что когда уменьшается проводимость Ks каналов, то фаза медленной деполяризация мембраны при ПД происходит медленнее и потенциал тоже медленее падает в минусовые значения, а значит Ca2+ каналы n-типа (которые являются высокопороговыми) остаются дольше открытыми и кальция больше вываливается в клетку. НО Я МОГУ БЫТЬ НЕ ПРАВА. Путь с 5 HT2 рецепторами ниже
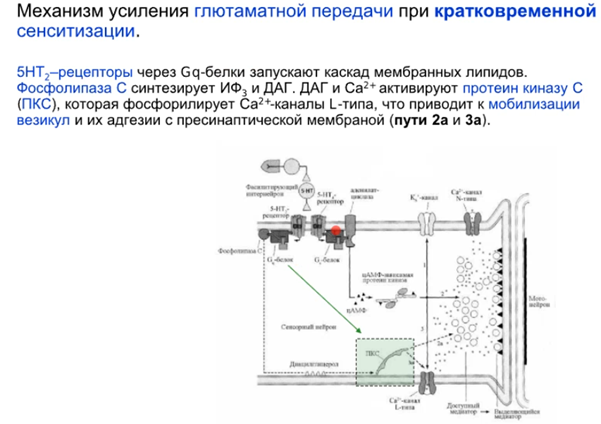
Комментарий (если кто забыл то Фосфолипаза С активируется субъединицами Gαq или Gβγ G-белка. Она гидролизует фосфатидилинозитол (PIP2) на два вторичных медиатора (IP3 и DAG), в описании пропущено это)
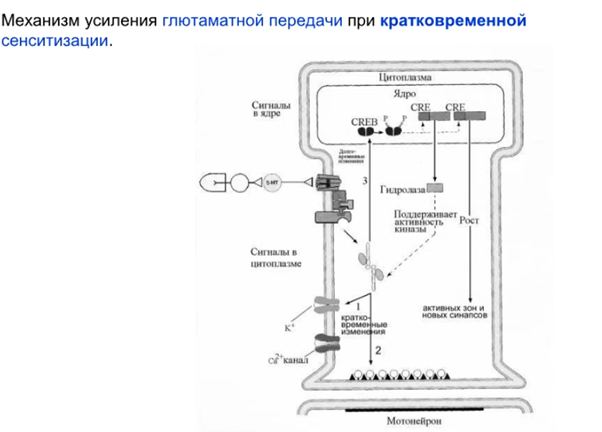
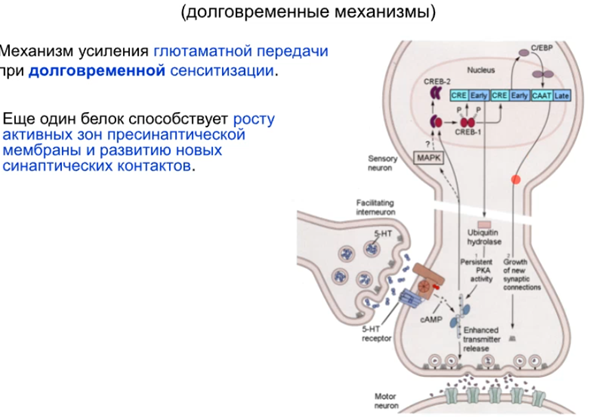
А сейчас рассмотрим более “продолжительную” кратковременную сенситизацию через ядро. Протеинкиназа А (PKA) проникает в ядро и активирует фактор транскрипции CREB и в результате (Латанов думает что мы все суперзнаем про транскрипцию и трансляцию, если че коротко тут можно в вики почитать в разделе механизм действия https://ru.wikipedia.org/wiki/CREB) получаем синтезированную гидролазу, которая блокирует регуляторные домены в РКА, что усиливает активность киназы (см ниже). Ну и таким образом я так понимаю PKA идет дальше фигачить фосфорилирование Ks каналов, что обеспечивает сенситизацию.
Ниже еще одна англояз картинка покрасивше если кому надо про серотониновые пути.
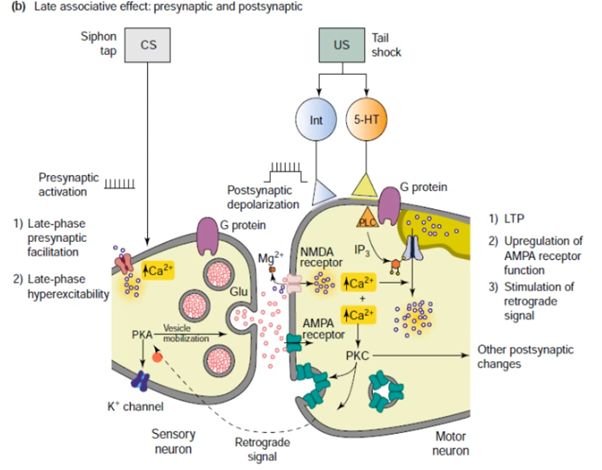
Убиквитин гидролаза на схеме выше относится к кратковременным эффектам. Из долговременных через ядро еще синтезируются(видимо через CAAT кусок ДНК на схеме) факторы роста, которые способствуют разрастанию синапса (может увеличится пресинаптич мембрана или отпочковаться коллатераль).
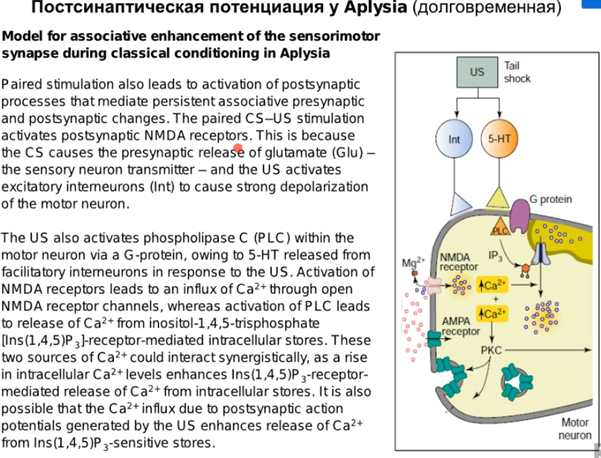
Постсинапс вносит свой вклад наравне с пресинапсом в этих работах, что мы ниже и рассмотрим.
исчо раз для тех кто не понял вся кратковременная хрень на англе повторить, А ТО ВДРУГ ЗАБЫЛИ.
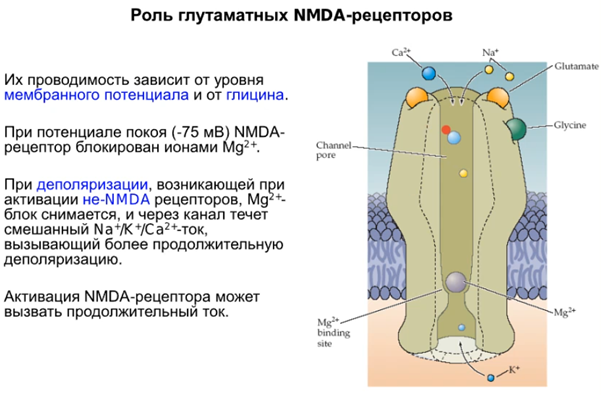
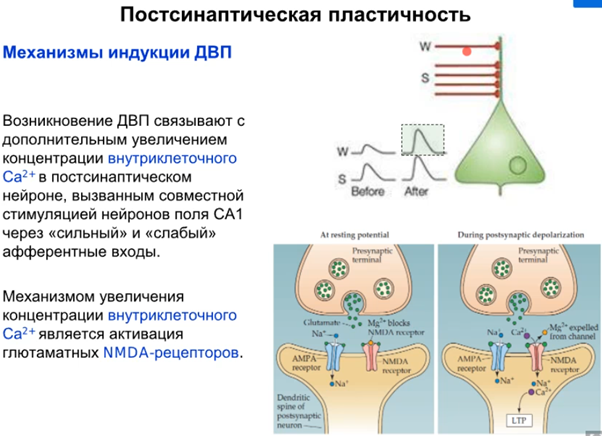
Наконец говорим про постсинапс уже, подробно на слайдах выше. Кратко от Латанова: изложение в терминах павловских представлений(ниже, CS - это условный стимул, US - безусловный). Парная стимуляция сифона и хвоста активирует NMDA - рецепторы(которые как мы знаем являются главным индуктором долговременной потенциации в гиппокампе). Посмотрим еще на серотонинэргический синапс(тут серотониновый рецептор 2 типа), активир. G белок, который активир. PLC (фосфолипаза С), которая разваливает PIP2 на IP3(вторичный мессенджер) и DAG. Когда в клетке увеличивается количество Ca2+, то его избыток закачивается в ЭПР переносчиками (которые потенциалзависимые и активируются внешний лигандом IP3, после чего Ca2+ вываливается из ЭПР во внутриклет. прос-во). На схеме нет, но в ЭПР еще есть рианодиновые рец-ры, которые активируются Ca2+, формируя таким образом петлю положительной обратной связи, начиная с переносчиков и волнами увеличивая конц. Ca 2+ внутри клетки. Второй источник повышенного содержания Ca2+ в клетке это AMDA рец-ры(которые обеспечивают деполяризацию для NMDA рец-ров, так как мы знаем что последние открываются только при наличии глутамата и деполяризации). При деполяризации Mg2+ из просвета канала уходит и NMDA рец-р долго пускает Ca2+ в клетку, что в свою очередь активирует PKC. Она активирует РАЗЛИЧНЫЕ процессы, в том числе передача сигнала в ядро для синтеза новых AMPA рец-ров и экзоцитоз этих AMPA рец-ров на мембрану, что приводит к усиленной активации моторного нейрона. Помимо этого происходит ретроградный сигнал в пресинапс на PKA для закрытия Ks каналов, что приводит к деполяризации.
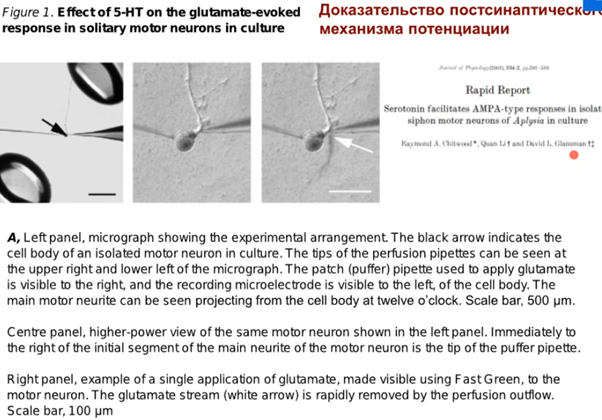
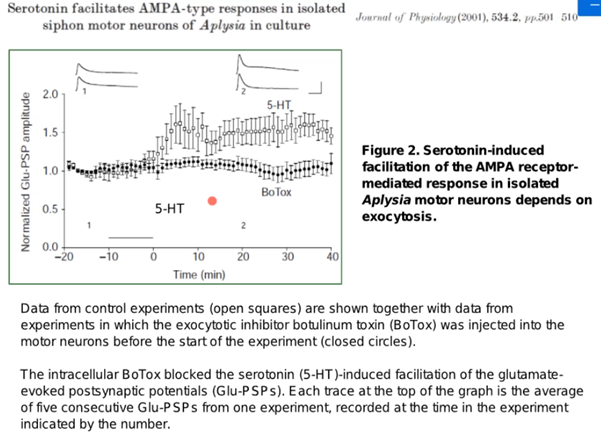
А теперь про эксперимент ниже, который произвел на Латанова неизгладимое впечатление про док-во постсинаптич. потенциации. Работа выполнена в тусе Глацмана, который аплизию истыкал уже бедняжку, на изолированном мотонейроне. На трех картинках из работ там видно слева на центр. картинке электрод в клетке и пипетку справа которая фигачит глутамат. Клетку помещали в разные среды и регистрировали электродом ВПСП, вызванный глутаматом, в варианте а (non facilitated glutamate response) сначала положили в изотонический раствор, а потом в р-рантагонист. Апликация антагониста(DNQX) AMPA рец-ров в р-ре немного ингибировала Глутаматный ВПСП в ответ на апликацию глутамата из пипетки, то есть немного выключили AMPA рецепторы.
Вариант (б). Сначала изотонический раствор, потом фигачнули глутамат на клетку и потом окунули на 10 минут в серотонин, а потом опять в изотонический раствор, то будет наблюдаться увеличение ВПСП в два раза. А после окунания в раствор с антагонистом наблюдаем резкое снижение этой пластичности на графике. Это говорит о том, что во время серотонина увеличилось количество AMPA рец-ров которые дали такой ответ и если мы устраняем антагонистом потом эту активность то и ВПСП тоже падает. При отмывке препарата все восстанавливается до 1 и даже чуть выше остается эффект от новых AMPA рец-ров.
Дальше(см выше) ребята решили не останавливаться и зафигачили еще ботокс, который как известно ингибирует процессы экзоцитоза,с помощью которого AMPA рец-ры встраиваются в мембрану. На графике в клетках, где был введен ботокс предварительно увеличения амплитуды ВПСП и не произошло после выливания на них серотониа, в клетках, где ботокса не было ВПСП увеличился соотвественно. Таким образом, было доказано, что серотонин участвует в процессах потенциации, что обусловлено встраиванием AMPA рец-ров в мембрану, поскольку блокирование их антагонистом устраняет эти эффекты. С ботоксом было показано, КАК это происходит, то есть путем экзоцитоза.
ЭТА ПРЕЗА ЗАКОНЧЕНА! НАЧИНАЕМ ДРУГУЮ на полчаса че успеем в нашей последнем лекции.
Рассмотрим механизмы постсинаптич. пластичности на гиппокампе, но сейчас коснемся молекулярных аспектов. Посмотрим какой раз на наш любимый NMDA рец-р и на слайде все понятно написано, добавлю только что магниевая пробка выходит из канала где-то при -30 - (-25) мВ при деполяризации и ток начинает течь.
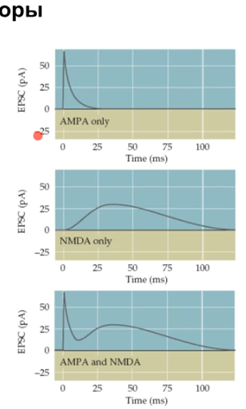
Ниже видно, что AMPA рец-ры вызывают быстрый, мощный, но кратковременный ток, а NMDA медленный, но продолжительный ток.
На слайде выше, где красная точка то же самое. Черная жирная линия ниже это активация только AMPA рец-ров, линия выше - вместе с NMDA.
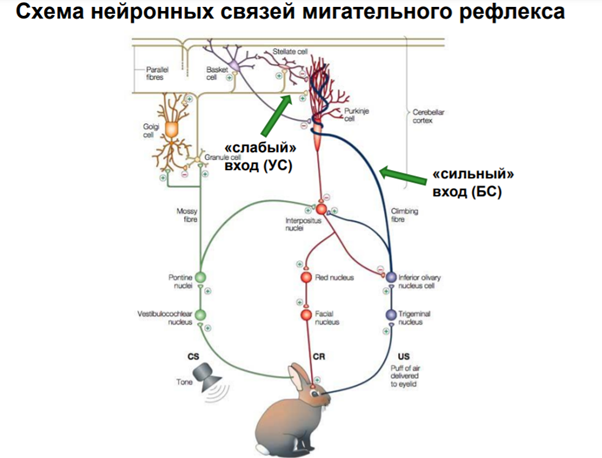
Активировали нейрон в поле СА1 через слабый вход(W) и сильный(S) вход. Потенциация чрез сильный вход дает в синапсе усиление ответа, через слабый ничего не давала. При одновременной стимуляции слабого и сильного позже произойдет потенциация и через слабый вход. Это объясняется включением nMDA рец-ров в процесс при сильной стимуляции и последующем массивном входе кальция, при слабом только AMPA рец-ры работают (некоторые редкие aMPA рец-ры могут помимо K и Na еще Ca пускать если есть спец. субъединица). Ниже опять какая то похожая картинка которые были выше про вклад AMPA и NMDA рец-ров
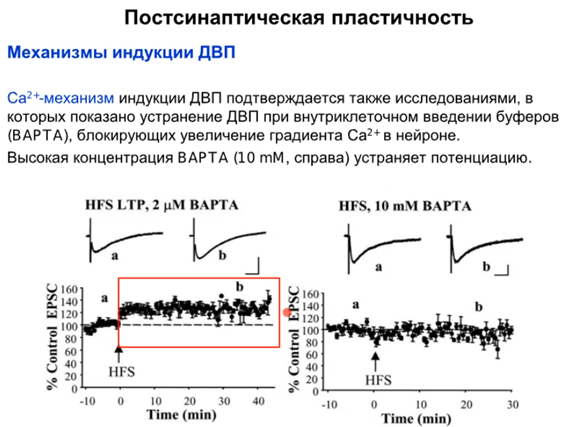
Что будет, если устранить экзоцитоз Ca2+(см ниже)? BAPTA является кальциевым буфером, который связывает кальций внутри клетки и он не выходит.На первом графике малые конц BAPTA при HFS(high frequency stimulation высокочастотная стимуляция) ничего не привносят в уменьшение потенциации, на втором большие дозы BAPTA уже устраняют потенциацию, ЗНАЧЕТ потенциация возможна только при участии кальция.
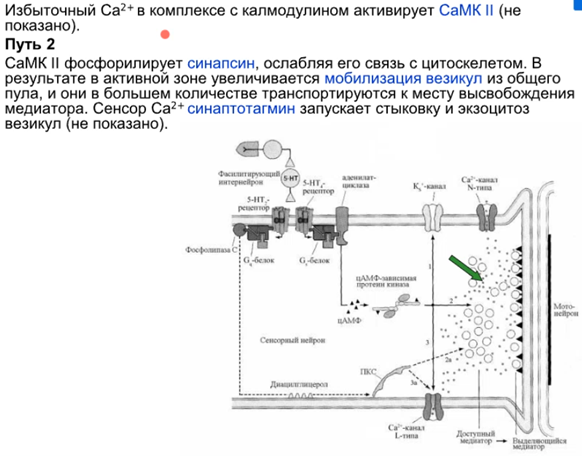
Еще кальций вываливается из внутриклеточных депо(ниже). Есть CaMKII(кальмодулин зависимая киназа) которая фосфорилирует НЕКИЕ субстраты, которые приводят к экзоцитозу AMPA рец-ров (на картинке просто нет каскада что конкретно фосфорилир, а Латанов видимо не помнит и думаю нам тоже не надо). Также важна PKA, которая синтезир. при цАМФ.
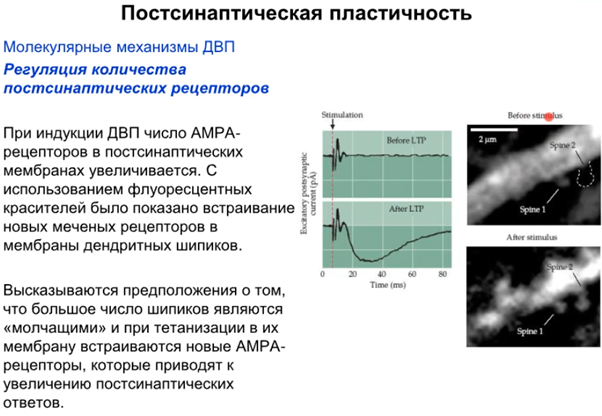
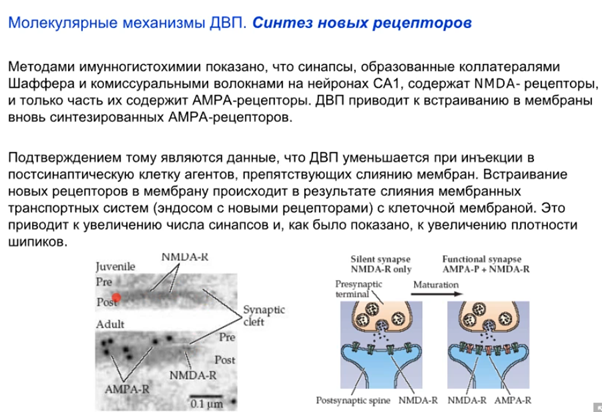
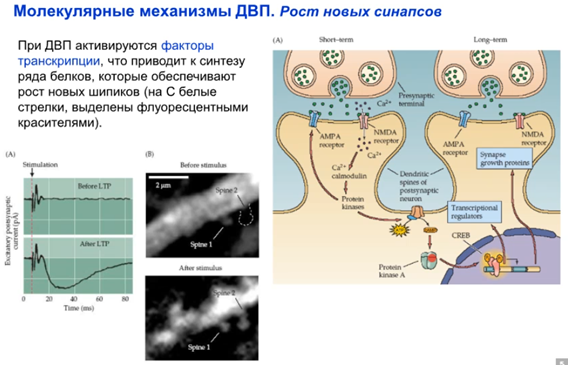
В работе ниже показано что при индукции ДВП AMPA рецепторы рили встраиваются и появляются новые синапсы. После стимуляции происходит разрастание синаптической поверхности в виде новых шипиков (spine 2 на картинке справа). Также говорится о том, что есть так называемые спящие синапсы, в которых на постсинапсе только NMDA рец-ры, а значит он не работает даже с глутаматом в щели (так как нужно до -30 мВ мембрану чем-то довести). Когда AMPA рец-ры встроились, синапс становится “говорящим”/функциональным. Еще третья картинка ниже там фотка молодого и созревшего синапса. В молодом там только NMDA а в зрелом еще и AMPA есть. значит он рабочий.
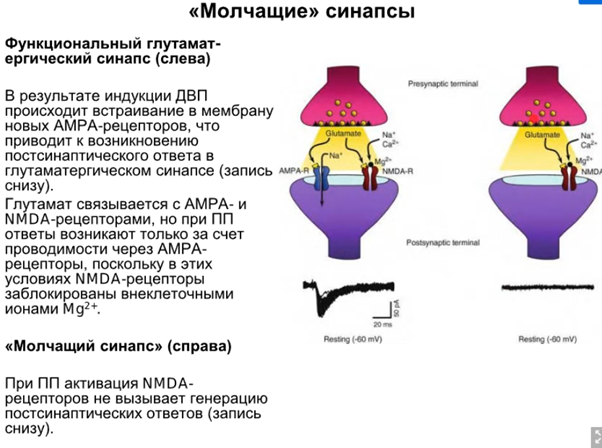
А ниже показали, что молчащие синапсы это РИЛИ NMDA рец-ры обеспечивают (на слайде все отлично расписано, Латанов почти читал сам). И еще ниже суперобощающая картинка, чтобы ТОЧНО запомнили.
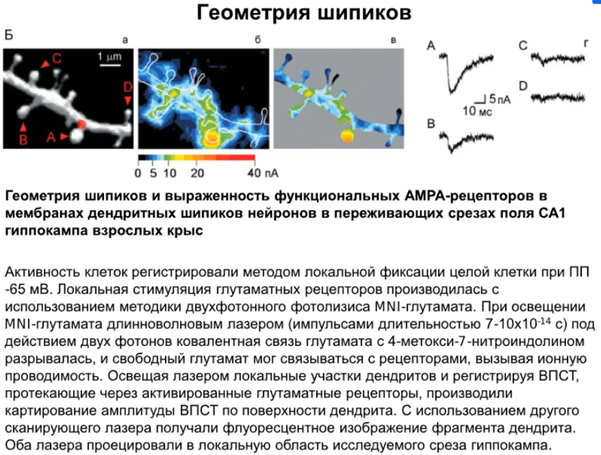
Про прикольный эскп про геометрию шипиков на последние 10 минут лекции, который мне понравился. На глутамат пришпандорили молекулу MNI (зеленая на слайде) с которой он не может связаться с AMPA рец-ром. Далее использовалась методика двухфотонного фотолизиса, где при освещении ЛУЧИКАМИ лазера глутамат освобождался от этой MNI в некоторых точках нашего дендритического объекта и одновременно картировали ВПСТ по поверхности дендрита. Там где синее, не оказалось особо AMPA рец-ров и эти шипики тонкие(или филоподии) с молчащими синапсами (C,D), а (A и B сидели на трубе) показали токи поактивнее и были грибовидными с норм количеством AMPA рец-ров. И типо все пластические перестройки могут идти только чрез грибовидные шипики получается.
Как далее происходит молекулярочкой разрастание синаптической поверхности? Это происходит через ядро с участием PKA и фактора транскрипции CREB, где синтезируются факторы роста для синапса (хотя на картинке написано белки роста синапса), и синаптическая поверхность разрастается и появляются новые шипики.
Еще раз че делает Ca2+ и CAMKII и короткое резюме про короткие и длит перестройки
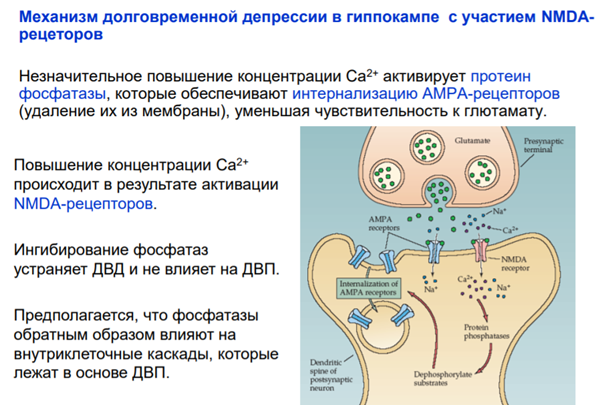
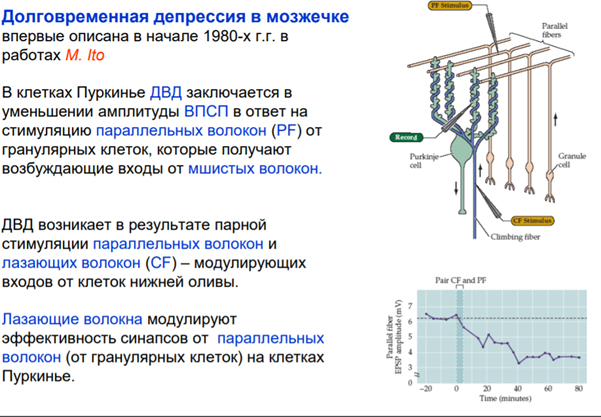
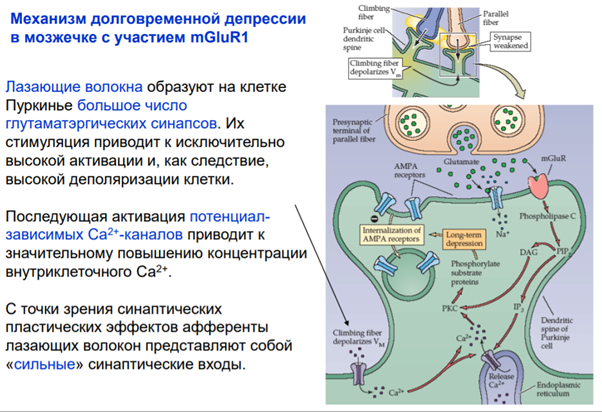
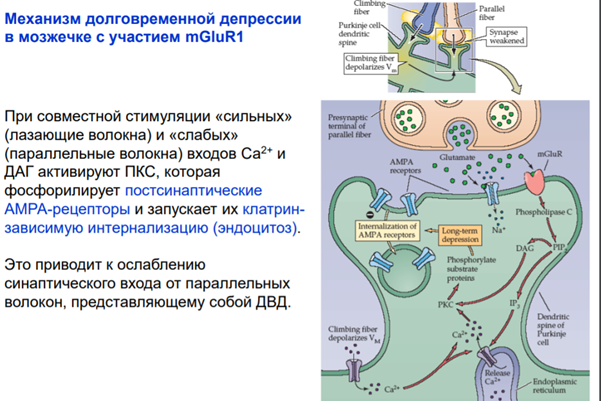
И еще один ФРАГМЕНТИК который Латанов сказал самим разобрать и я это делаю. И это про постсинаптическую депрессю. Там вроде все понятно просто приложу слайды
|
|||||||||
|
|
Последнее изменение этой страницы: 2022-01-22; просмотров: 65; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.119.172.146 (0.072 с.) |

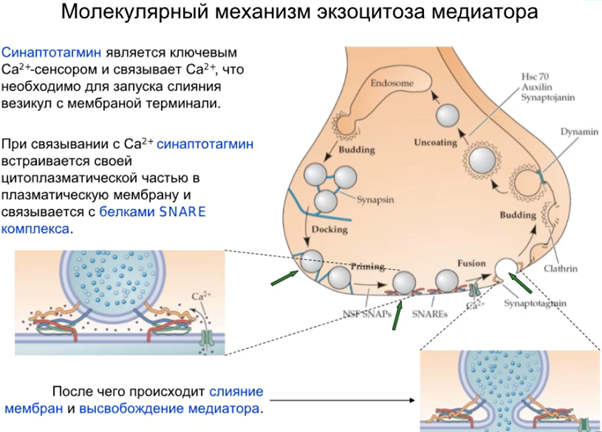
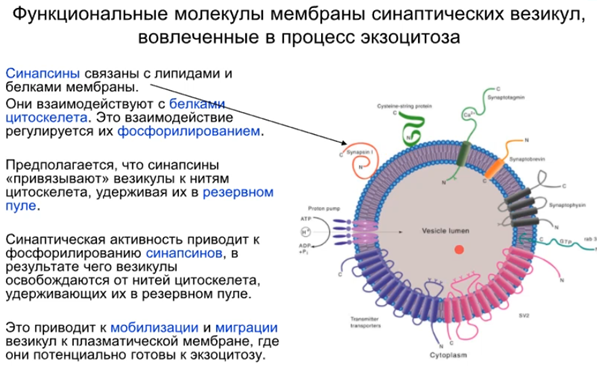
 Синапсин держит везикулы в неком пуле, они вместе связаны (предыдущая картинка). В нефосфорилированном состоянии синапсины присоединяются к синаптическим пузырькам и запускают полимеризацию актина, а фосфорилирование вызывает их отсоединение от пузырьков.
Синапсин держит везикулы в неком пуле, они вместе связаны (предыдущая картинка). В нефосфорилированном состоянии синапсины присоединяются к синаптическим пузырькам и запускают полимеризацию актина, а фосфорилирование вызывает их отсоединение от пузырьков.