Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Нейрохимические механизмы пластичности. Обучение
Следующий раздел - нейрохимические механизмы пластичности нейрона на клеточных аналогах обучения. Та же самая парадигма, та же самая депрессия. Мы очень подробно разобрали влияние транспортных систем на мобильность рецепторов, на пластичность, а теперь, собственно, переходим к нейрохимическому разделу.
В чем заключаются нейрохимические механизмы - это химическая модификация структурных компонентов рецепторов и ионных каналов плазматической мембраны нейрона (это относительно подробно разобрали на предыдущих лекциях) и внеклеточные и внутриклеточные сигнальные молекулы (первичные,вторичные, третичные посредники в нейроне). Речь пойдет в основном про метаботропные каскады.
• деполяризация терминали • первичный химический посредник (медиатор, внеклеточная среда) • активация постсинаптического рецептора • вторичный посредник (цитоплазма) • активация протеинкиназы и ее транслокация в ядро (протеинкиназа может дать сигнал в ядро, протеинкиназа а, например, проникает в ядро) • третичный посредник (ядро) (фактор транскрипции) • последовательная активация ранних и поздних генов Это самая общая схема, не всегда доходит до транскрипции, но чтобы обеспечить долговременные процессы, это нужно. При кратковременной пластичности могут быть, наверное, активированы ранние гены. И в конечном счете ответ - реакция. Реакция заключается в том, что нейрон демонстрирует тот или иной постсинаптический потенциал. А если это командный нейрон, то он обеспечивает соответствующие поведенческие реакции.
Много вторичных посредников есть. Пивоваров сказал, что их 49. Вторичные посредники образуются из того, что есть, из доступных субстратов и имеют относительно короткий биохимический полупериод (полупериод - время, за которое содержание того или иного вещества уменьшается в два раза), полупериод их утилизации. Самые известные известные вторичные посредники - цАМФ и цГМФ. Остальное на слайде.
Лекотриены, гепоксилины, липоклины - липидные посредники. И если арахидоновая кислота может быть, например, вторичным посредником, то ее производные уже третичные и они превращаются в четвертичные и так далее.
Вторичным мессенджером считают еще цАМФ и кальций, происхождение которых не в результате синтеза и так далее, а в результате работы Na+,K+-помпы.
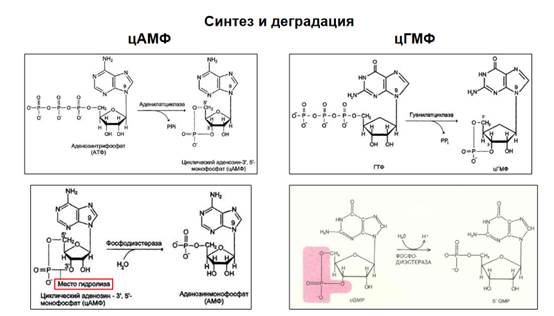
Биосинтез ясен - цАМФ и цГМФ синтезируются аденилатциклазами, а утилизируются фосфодиэстеразами, которые превращают их в линейную 5’3’форму. Комлекс кальций-кальмодулин много что делает, например, активирует протеинкиназу. Кальций-кальмодулин-активируемые киназы 4 типа хорошо известны.
Напоминание, как выглядит синтез и деградация цАМФ и цГМФ. цАМФ получается из АТФ путем отщепления двух фосфатов и образования эфирной связи между фрагментом пятичленного цикла через кислород на метил того же цикла. Фосфодиэстераза гидролизует эту эфирную связь и остается только 5’3’аденозинмонофосфат линейный, не циклический. То же самое в ГМФ.
Эти вторичные посредники активируют свои протеинкиназы. цАМФ активирует протеинкиназу а, а цГМФ - протеинкиназу G. Протеинкиназы имеют регуляторные субъединицы и каталитические субъединицы. Каталитические субъединицы контролируются регуляторными. Для того, чтобы протеинкиназа заработала необходимо, чтобы 4 молекулы цАМФ активировали регуляторные субъединицы. В результате взаимодействия со вторичным посредником каталитические субъединицы освобождаются от супрессии со стороны регуляторных субъединиц и выполняют свою роль. При уменьшении цАМФ активируется меньше протеинкиназ, а это уменьшение происходит, если мы блокируем работу циклаз соответствующих.
Уровень кальция внутри клетки очень низкий - 10-100 нМ. Поддерживается это кальциевыми АТФ-азами (кальциевыми насосами). Кальциевые насосы удаляют кальций из клетки и удаляют его в окружающую среду - это кальциевые помпы в плазматических мембранах, есть еще кальциевые помпы в мембранах эндоплазматического ретикулума, и кальций закачивается в эти мембраны, в эти внутриклеточные органеллы - внутриклеточные цистерны - с помощью специализированной помпы. Есть еще кальций-связывающие белки - калбидины, он фиксирует растворенный кальций и препятствует его участию в каскадах. Есть еще натрий-кальциевый обменник, который очень мощный механизм, может существенно повлиять на потенциал мембраны.
При активации иногда происходит резкое повышение концентрации кальция в цитоплазме иногда на два-три порядка. Открываются потенциал-зависимые кальциевые каналы на плазматической мембране, если это синаптическое воздействие какое-то или электрическое воздействие. Так же кальций может поступить из внутриклеточного депо - из ЭПР. На слайде написано про то, что нейромедиаторы типа глутамата и АТФ активируют ионотропные рецепторы, которые пропускают кальций. АТФ рецепторы вообще не селективны к катионам, глутаматные (в основном, AMPA) при потенциале покоя активируются, кальций тоже проходит по AMPA-рецепторам. Извлечение кальция из внутриклеточных органелл происходит с участием вторичных посредников - инозитолтрифосфат и цАМФ, а также рианодин тоже вызывает высвобождение кальция. Иногда кальций может выходить из буферов,то есть выходить из фиксированного состояния, отщепляясь от буфера.
Кальций, в общем, ключевой элемент, который запускает много различных процессов. Если кальция очень много, то будет цитотоксическое действие - большая кальциевая нагрузка внутри клетки может вызвать ее гибель. Это используют при воздействие на NMDA-рецепторы - если они долго будут активированы, то клетка погибнет из-за того, что в результате каскадов повышается концентрация активных форм кислорода.
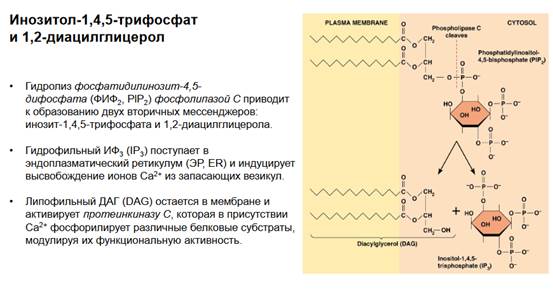
Мембранное происхождение имеют два вторичных посредника - это инозитолтрифосфат и диацилглицерол. Инозитолтрифосфат получается из ФИФ2 (фосфатидилинозит-4,5-дифосфат), фосфолипаза С разлагает ФИФ2 на инозитолтрифосфат и диацилглицерол. Путем разрезания фосфолипазой С связи, которая показана на рисунке красной пунктирной линией, отваливается инозитолтрифосфат и диацилглицерол. Поскольку диацилглицерол - это органический спирт, он связан с мембраной липофильными концами (на рисунке показан его хвост), он остается в мембране, то есть в дальнейшем активирует протеинкиназу С, которая так же ассоциирована с мембраной. А инозитолтрифосфат в свободном виде вываливается в цитоплазму, он растворим и, соответственно, он может привести к высвобождению кальция из внутриклеточных везикул, поскольку на мембранах этих внутриклеточных везикул есть специальные рецепторы, которые активируются инозитолтрифосфатом, эти рецепторы структурно очень напоминают потенциал-зависимые каналы - имеют 6 трансмембранных сегментов, водную пору и активируются этим лигандом.
Аденилатциклаза, фосфодиэстераза и протеинкиназа а обеспечивают каскад, связанный с действием цАМФ. Активация G-белка какого-нибудь рецептора, который активируется различными нейромедиаторами (например, тем же самым ацетилхолином, норадреналином), этот G-белок активирует аденилатциклазу, дальше получается цАМФ, который активирует протеинкиназу и дальше много эффектов. Также цАМФ может воздействовать на ионный канал. Показаны еще некоторые белки, которые могут приводить к синтезу цАМФ.
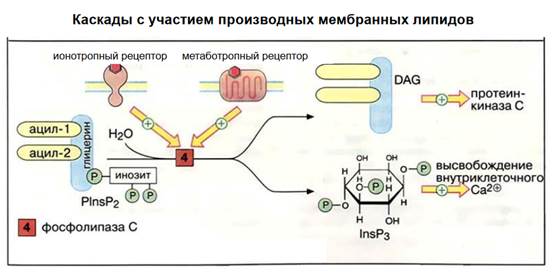
Это мембранные липиды. Вот там красненький метаботропный рецептор. Фосфолипид распадается на диацилглицерол и инозитолтрифосфат. Эти вторичные посредники имеют свои каскады. В конечном счете активируется фосфолипаза С с участием диацилглицерола, а инозитолтрифосфат увеличивает концентрацию кальция и кальций в данном случае будет уже третичным посредником.
Показаны каскады с участием кальция, они многообразны.
Дальше ретроградные вторичные посредники - это NO, арахидоновая кислота и ациклические эйкозаноиды, еще эндоканабиноиды. Красной рамки - пример того, как получается NO - при действии на NMDA-рецептор может запуститься NO-синтаза и из L-аргинина образуется NO. Сам NO газ и разлетается на несколько микрон вокруг того места, откуда он выскочил. И этот NO может цепляться на молекулы везикул и нарушать экзоцитоз из этих везикул и может так же воздействовать на полтенциал-зависимые кальциевые каналы, которые обеспечивают, собственно, этот экзоцитоз. Ретроградная регуляция широко используется для нормализации работы синапсов.
|
|||||||
|
|
Последнее изменение этой страницы: 2022-01-22; просмотров: 38; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.189.2.122 (0.011 с.) |


 Последовательность процессов при кратковременной пластичности. Депрессия - это кратковременная пластичность.
Последовательность процессов при кратковременной пластичности. Депрессия - это кратковременная пластичность.