Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Роль серотонина в пластичности нейрона на клеточном аналоге обучения
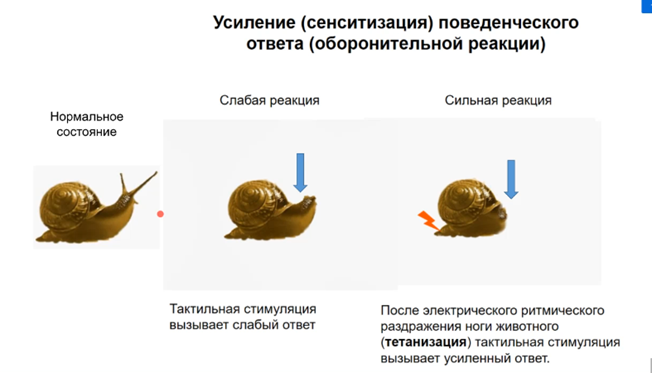
Мы разберем роль ключевого медиатора, который обеспечивает собственно основные перестройки в пресинаптической и в постсинаптической пластичности. Это избегательная реакция на моллюсках – аплизии и улиток. Напоминаю Вам с чего начались исследования Пивоварова. Вот эти работы (усиление – сенситизация поведенческого ответа оборонительной реакции) мы рассмотрели в первых лекциях. Оборонительная реакция заключалась в том, что если коснуться антенн, то улиточка сжимается, но сжимается не очень сильно. Перед тем как тестировать такую оборонительную реакцию дают ритмический удар по хвосту, который вызывает тетанизацию, если потом улиточку стимулировать, то она будет давать больший поведенческий ответ.
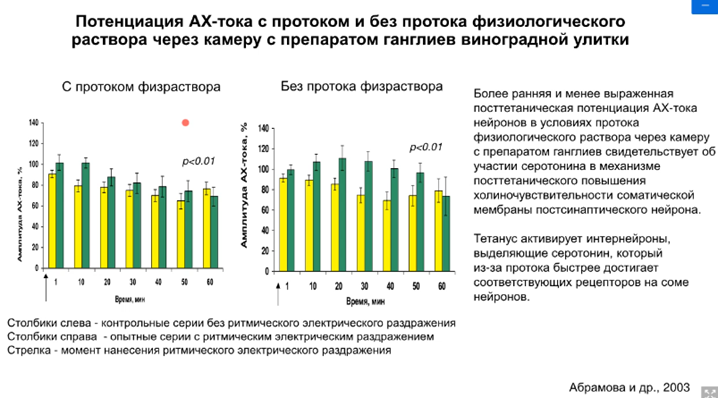
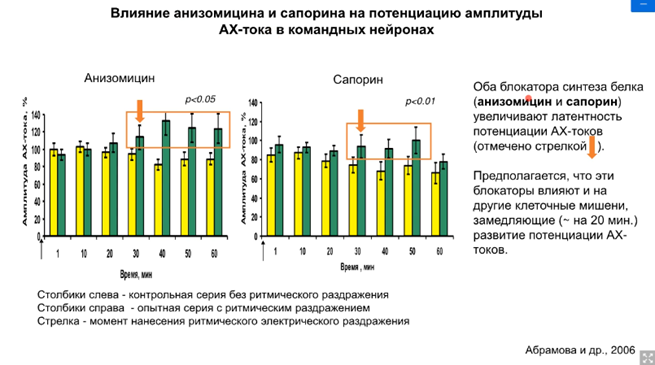
А на токах мы увидим (это делали уже на клеточной модели). Можем увидеть следующую динамику токов:
Желтое – контроль. Это искуственный синапс. Апликация АЦХ на поверхность командного нейрона. Измеряется внутриклеточный ток командного нейрона, наблюдаем относительно умеренную депрессию. Потом даем сенсетизирующий стимул и наносим аппликацию АЦХ. Сенситизация ранняя, обратите внимание (с протоком физраствора). Без протока (наш препарат пребывает в несвежем растворе. Было высказано предположение, что в нашем растворе может накапливаться некая субстанция, которая и обеспечивает олее продолжительные эффекты, только на второй стимул, а продолжается эффект дольше). Объясняется это тем, что в межклеточной среде накапливается серотонин, который обеспечивает такую холинчувствительность мембраны по синаптическому нейрону. Предположили, что тетанус активирует интернейроны, которые выделяют серотонин, который из-за протока быстрее достигает рецепторов и начинается быстрее эффект, но непродолжительный.
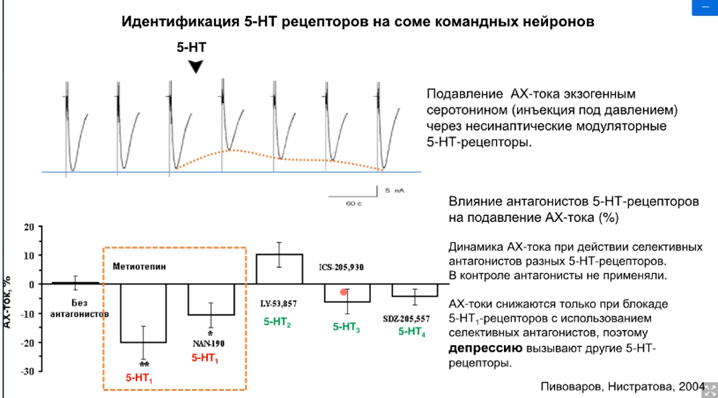
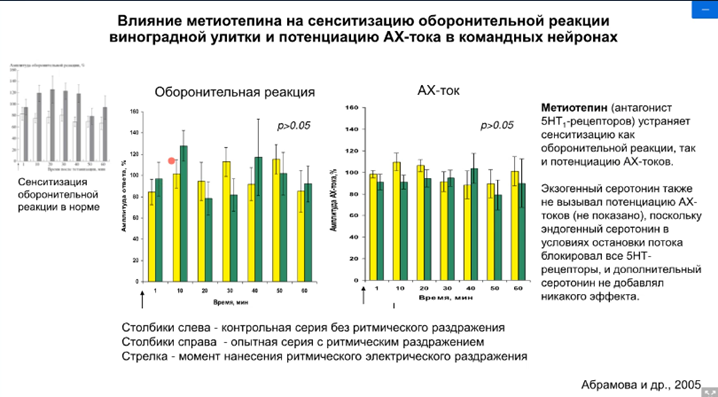
Оставалось выяснить, какие же рецепторы. Было проверено на депрессии. Применяли стимуляцию АЦХ, а потом применили серотонин, но не стимулировали тетанусом входы серотонина. В эксперименте использовали специфические блокаторы серотонина 5-НТ1, 5-НТ2, 5-НТ3 иноторопный, 5-НТ4. Оказалось, что депрессия сохраняется при блокаде 5-НТ1, это означает, что не участвуют в пластических перестройках. На самом деле это была проверка результатов, полученных другими учеными, которые работали до Пивоварова на аплизиях – это работа группа Кендала и Глазмана (там речь шла о рецепторах 2 и 4 типа; они были достаточно хорошо изучены).
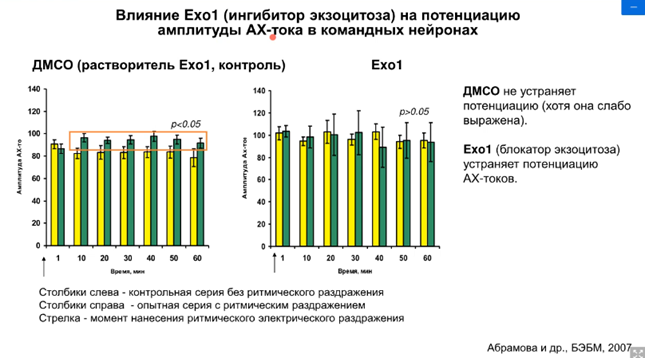
Динамин участвует в обратном захват рецепторов из мембраны. Он образует замкнутые эндосомы, которые захватают рецепторы из мембраны. Т.е. две функции – интернализация и рецклирование. Рециклирование необходимо для обеспечения сенсатизации.
Аналогичный эффект нужно посмотреть и у аплизии. Это в следующей лекции
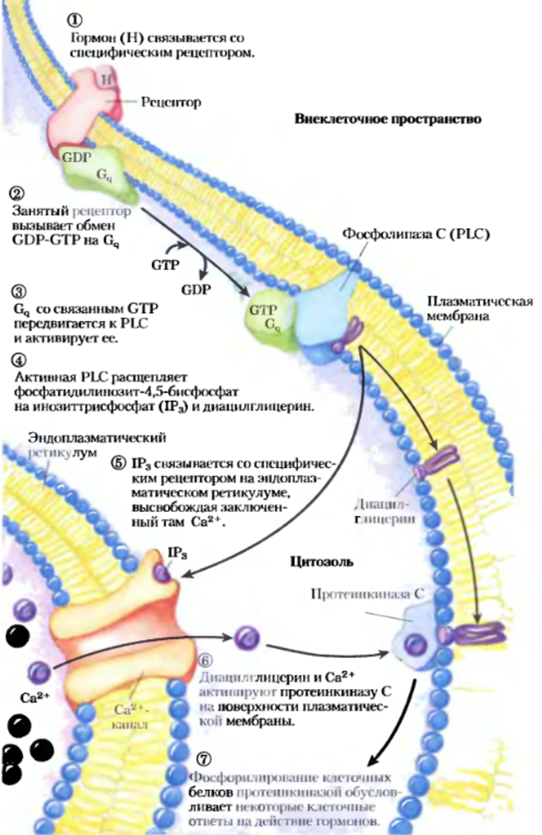
Другими словами, что хотел сказать Латанов по биохимии: Диацилглицерин, инозиттрисфосфат и Са2+ играют сходные роли вторичных мессенджеров. Еще один большой класс GPCR — рецепторы, через G-белки сопряженные с фосфолипазой С (PLC) плазматической мембраны, которая специфична в отношении содержащегося в плазматической мембране липида фосфатидилинозит- 4,5-бисфосфата. Когда гормон этого класса связывается с соответствующим специфическим рецептором на плазматической мембране (стадия 1), комплекс гормон-рецептор катализирует обмен GTP-GDP на ассоциированном G-белкс Gq (стадия 2), активируя его практически так же, как (3-адренсргический рецептор активирует Gs). Активированный G4 в свою очередь активирует специфичную для PIP2 PLC (стадия 3), которая катализирует (стадия 4) образование двух мощных вторичных мессенджеров: диацилглицерина и инозит-1,4,5-трисфосфата, или IР3 (не путать с Р1Р3, смотри в конце). Растворимый в воде инозиттрисфосфат диффундирует из плазматической мембраны в эндоплазматический ретикулум, где связывается со специфическими рецепторами IР3 и заставляет открыться Са2+-каналы, находящиеся в ЭР. Вследствие этого в цитозоль высвобождается Са2+ (стадия 5), концентрация Са2+ в цитоплазме резко возрастает до ~10-6 М. Одним из эффектов увеличения концентрации Са2+ является активация протеинкиназы С (РКС). При активации РКС диацилглицерин кооперируется с Са2+, также действуя как вторичный мсссснджер (стадия 6). РКС фосфорилирует остатки Ser или Thr специфических белков-мишеней, изменяя их каталитическую активность (стадия 7).
Существует целый ряд изоферментов РКС, каждый имеет характерное распределение по тканям, специфичность к белкам-мишеням и различные функции в клетке. Среди мишеней — белки цитоскелета, ферменты и ядерные белки, регулирующие экспрессию генов. Все это семейство ферментов отличается широким спектром активностей, например они влияют на процессы в нервной и иммунной системах, а также регулируют деление клеток.
|
||||||
|
|
Последнее изменение этой страницы: 2022-01-22; просмотров: 39; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.30.162 (0.007 с.) |